Лето круглый год: Мадейра готова принимать россиян
https://ria.ru/20210607/madeyra-1735459439.html
Лето круглый год: Мадейра готова принимать россиян
Лето круглый год: Мадейра готова принимать россиян — РИА Новости, 07.06.2021
Лето круглый год: Мадейра готова принимать россиян
Обладатели действующей шенгенской визы могут отправиться на португальскую Мадейру — архипелаг, удаленный от континентальной части страны на тысячу километров… РИА Новости, 07.06.2021
2021-06-07T08:00
2021-06-07T08:00
2021-06-07T08:05
туризм
достопримечательности
португалия
мадейра
туризм
куда поехать
куда можно лететь
/html/head/meta[@name=’og:title’]/@content
/html/head/meta[@name=’og:description’]/@content
https://cdnn21.img.ria.ru/images/07e5/06/03/1735471435_0:0:3639:2047_1920x0_80_0_0_3a1e6cfa90448389f4248772932076e2.jpg
МОСКВА, 7 июн — РИА Новости, Мария Селиванова. Обладатели действующей шенгенской визы могут отправиться на португальскую Мадейру — архипелаг, удаленный от континентальной части страны на тысячу километров. Остров открыл границы для российских туристов еще в конце апреля, правда, добраться до него можно только с пересадкой. Зато там лето круглый год, 150 километров пляжей, необычные леса, колоритные деревушки, вкусные рыба и вино. Как долететь до Мадейры, что там смотреть, есть и пить — в фотоленте РИА Новости.
Остров открыл границы для российских туристов еще в конце апреля, правда, добраться до него можно только с пересадкой. Зато там лето круглый год, 150 километров пляжей, необычные леса, колоритные деревушки, вкусные рыба и вино. Как долететь до Мадейры, что там смотреть, есть и пить — в фотоленте РИА Новости.
португалия
мадейра
РИА Новости
7 495 645-6601
ФГУП МИА «Россия сегодня»
https://xn--c1acbl2abdlkab1og.xn--p1ai/awards/
2021
РИА Новости
7 495 645-6601
ФГУП МИА «Россия сегодня»
https://xn--c1acbl2abdlkab1og.xn--p1ai/awards/
Новости
ru-RU
https://ria.ru/docs/about/copyright.html
https://xn--c1acbl2abdlkab1og.xn--p1ai/
РИА Новости
7 495 645-6601
ФГУП МИА «Россия сегодня»
https://xn--c1acbl2abdlkab1og.xn--p1ai/awards/
https://cdnn21.img. ria.ru/images/07e5/06/03/1735471435_0:0:2729:2047_1920x0_80_0_0_825e11e3c6ef71a864aafb9a938d0ef1.jpg
ria.ru/images/07e5/06/03/1735471435_0:0:2729:2047_1920x0_80_0_0_825e11e3c6ef71a864aafb9a938d0ef1.jpgРИА Новости
7 495 645-6601
ФГУП МИА «Россия сегодня»
https://xn--c1acbl2abdlkab1og.xn--p1ai/awards/
РИА Новости
7 495 645-6601
ФГУП МИА «Россия сегодня»
https://xn--c1acbl2abdlkab1og.xn--p1ai/awards/
достопримечательности, португалия, мадейра, туризм, куда поехать, куда можно лететь, фото, еда, рецепты, что приготовить, отдых на море, где отдохнуть
Сколько островов у архипелага Мадейра?
Обычно многие посетители острова, даже те, кто видел его давно, не знают, сколько островов на архипелаге Мадейра.
Архипелаг Мадейра расположен в Атлантическом океане между 30 ° и 33 ° северной широты, в 978 км к юго-западу от Лиссабона и примерно в 700 км от побережья Африки, почти на той же широте, что и Касабланка, относительно недалеко от Гибралтарского пролива.
Хотя обычно он считается европейской территорией, географически архипелаг уже расположен на континентальном шельфе Африки. Ближайший морской сосед — испанский архипелаг Канарских островов.
Сколько островов у архипелага Мадейра?
1. Остров Мадейра
Острова архипелага Мадейра: остров Мадейра — изображение 2
Остров Мадейра — главный остров архипелага Мадейра, расположенный в Атлантическом океане, к юго-западу от побережья Португалии, присоединенный к Европейскому Союзу. Вместе с Порту-Санту, необитаемыми островами и островами Сельвагенс это архипелаг Мадейра и автономный регион Мадейра, столицей которого является город Фуншал.
Остров Мадейра имеет вулканическое происхождение, площадью 742.4 км², обширную эндемичную флору, местную и экзотическую, и его типичный средиземноморский климат в основном находится на южной стороне, переходя в умеренный климат с более высокими квотами. Экономика широко ориентирована на туризм.
2. Остров Порто Санто
Острова архипелага Мадейра: остров Порту-Санту — изображение 3
Порту-Санту — португальский муниципалитет, который занимает весь остров Порту-Санту, автономный регион Мадейра, расположенный в деревне с названием, отличным от названия муниципалитета, городе Вила-Балейра. Его площадь составляет 42.48 км².
Его площадь составляет 42.48 км².
Это один из шести муниципалитетов Португалии, в которых есть один приход с таким же названием. Ближайший муниципалитет — Машику на острове Мадейра, расположенный к юго-западу. Мадейранцы часто называют его Золотым островом.
3. Пустынные острова
Острова архипелага Мадейра: необитаемые острова — изображение 4
Острова Дезерташ — это субархипелаг архипелага Мадейра, Португалия, имеющий вулканическое происхождение, расположенный к юго-востоку от острова Мадейра. Это природный заповедник Пустынных островов, также классифицированный Советом Европы как биогенетический заповедник.
Они являются частью Desertislands, Islet Floor, Deserta Grande и Bugio. В административном отношении они являются частью прихода Санта-Крус, муниципалитет Санта-Крус, автономный регион Мадейра.
4. Дикие острова
Острова архипелага Мадейра: дикие острова — изображение 5
Острова Сельвагенс — это субархипелаг архипелага Мадейра в Португалии, расположенный в Атлантическом океане. В административном отношении они являются частью прихода Се, муниципия Фуншал, Автономный регион Мадейра.
В административном отношении они являются частью прихода Се, муниципия Фуншал, Автономный регион Мадейра.
Они находятся в 165 км к северу от архипелага Канарских островов, в 250 км к югу от города Фуншал (Мадейра), примерно в 250 км к западу от побережья Африки, примерно в 1000 км к юго-западу от европейского континента.
Дикари состоят из двух основных островов и нескольких островков, которые, как и почти все острова Макаронезии, имеют вулканическое происхождение. Архипелаг — заповедник птиц, он очень суровый и имеет общую площадь 273 гектара.
Вы хотите свободы передвижения на острове Мадейра?
Лучший способ передвигаться по острову Мадейра — на одном из наших автомобилей. В 7М Прокат автомобилей Вы можете найти идеальный автомобиль для передвижения во время отпуска на Мадейре. Арендуйте любой из наших самых дешевых или роскошных автомобилей.
Карта острова Мадейра, цены на 2021-2022 год
Компания «Свои люди» дарит вам возможность посетить один из лучших уголков Европы. Карта острова Мадейра поможет вам получить хотя бы первое, поверхностное впечатление об этом замечательном месте с прекрасной природой и климатом.
Карта острова Мадейра поможет вам получить хотя бы первое, поверхностное впечатление об этом замечательном месте с прекрасной природой и климатом.
Карта острова Мадейра
Остров Мадейра принадлежит Португалии как автономный регион. Глядя на карту острова Мадейра, мы можем увидеть, что архипелаг состоит из двух густонаселенных островов: Мадейры и Порту-Санту, а также двух несколько отдаленных групп мелких необитаемых островов, которые служат пристанищем для морских птиц.
Карта острова Мадейра
Остров Мадейра на карте
Мадейра — португальский архипелаг и остров, который расположен в северной части Атлантики. Мадейра находится примерно в 1000 километров к югу от самой Португалии и приблизительно в 500 километров к западу от побережья Африки.
На карте острова Мадейра выделяется его столица Фуншал — город который главенствует над южным побережьем главного острова архипелага. Другими городами, с которыми нас знакомит карта острова Мадейра, являются: Машику, Камара-де-Лобуш, Вила-Балейра, Санта-Круш, Канису и Сантана.
Фуншал — остров Мадейра, Португалия
Мадейра представляет собой крупнейший остров архипелага общей площадью свыше 740 км², его длина – 57 км, а ширина – 22 км, общая длина побережья – 150 км. Рассматривая остров Мадейра на карте, видим, что длиннейшая ось находится между западом и востоком. Именно вдоль нее лежит цепь гор, высота которых доходит до 1220 метров. Самая высокая точка Мадейры — Пику-Руйву — достигает 1862 метров над уровнем моря.
Стоит отметить, что на юге осталось совсем мало субтропических лесов, а раньше они покрывали практически весь остров и даже дали ему название: по-португальски слово «мадейра» означает «древесина».
Мадейра является очень популярным круглогодичным местом отдыха. В первую очередь он знаменит вином здешнего производства под названием «мадера», а также прекрасными цветами и новогодними празднованиями с потрясающим фейерверком, который, по версии Книги Гиннесса, является самым большим в мире.
Мадейра (архипелаг)
Все, кто хоть раз бывали на Мадейре, остаются влюбленными в это волшебное место всю жизнь. Здесь горы находятся просто на берегу океана, и они полны жизни. С самых вершин весело бегут горные ручьи и шумят каскады водопадов, многие из которых образуют небольшие горные озера-бассейны. А вокруг буйство зелени, разбавленное яркими красками и благоуханием экзотических цветов.
Здесь горы находятся просто на берегу океана, и они полны жизни. С самых вершин весело бегут горные ручьи и шумят каскады водопадов, многие из которых образуют небольшие горные озера-бассейны. А вокруг буйство зелени, разбавленное яркими красками и благоуханием экзотических цветов.
Даже зимой температура воздуха здесь не опускается ниже +18 °С, тогда как летом никогда не бывает знойно и столбик термометра держится у отметки +26 °С. И даже прибережные воды Мадейры всегда теплые, ведь совсем рядом проходит Гольфстрим.
Мадейра
Открыт был остров в 15 веке Жоао Зарком, корабль которого был прибит к берегам Мадейры штормом. Он и дал название острову, которое переводится как «лес». Сегодня леса Мадейры являются заповедными и занимают более шестидесяти процентов всей территории. Не меньшее значение имеют и ботанические сады, где собраны экзотические растения со всего мира. А если вы захотите увидеть все это великолепие с высоты птичьего полета, то можно совершить восхождение на Пику-Руйву, являющуюся самой высокой точкой острова, отсюда вся Мадейра — как на ладони.
Стоит отметить, что на самом острове нет пляжей, они находятся на близлежащем островке Порту-Санту. А так, каждый отель располагает огромным плавательным бассейном. Вообще отели Мадейры отличаются высоким уровнем сервиса, практически в каждом из них есть SPA-центры, где осуществляются лучшие в мире программы по уходу за кожей лица и тела. Большинство отелей четырех- и пятизвездочные. Здесь есть все для активного отдыха — спортивные площадки, теннисные корты, горные веломаршруты, огромный аквапарк и т. д.
Порту-Санту
Насколько прекрасна природа Мадейры, настолько интересно ее архитектурное наследие. Чего только стоит старинный женский монастырь в окрестностях небольшого селенья Куррал-даш-Фрейаш. Здесь, когда сотни лет назад пираты регулярно совершали вылазки на остров, за высокими неприступными стенами монахини чувствовали себя в безопасности. Узнать о быте потомственных рыбаков Мадейры, и посмотреть, как готовят, а главное попробовать, знаменитую на весь мир мадеру можно в деревушке Камара-до-Лобуш. В Фуншале находится великолепный собор Святой Клары и старинная фабрика, на которой производится знаменитая вышивка «бурдадош».
В Фуншале находится великолепный собор Святой Клары и старинная фабрика, на которой производится знаменитая вышивка «бурдадош».
Прежде всего, на Мадейре стоит попробовать «Espetada de Carne» — запеченное на костре мясо, маринованное в особом соусе, приготовленную по традиционным рецептом рыбу-шпагу, ну и конечно же экзотические фрукты, которые растут только здесь.
Где остановиться: В качестве места для отдыха архипелаг Мадейра лучше всего подойдёт искателям полного релакса без упора на пляжи: таковых здесь нет по причине скалистой береговой линии, зато талассоцентров – хоть отбавляй. Из прочих бонусов – красивейшая природа и тишина. Главный город острова Мадейра Фуншал порадует более-менее удобными обустроенными пляжами и массой достопримечательностей. А вот на Порту-Санту памятников почти нет – основные туристы здесь – любители тишины и поклонники активных видов водного спорта.
Что посмотреть: Природные достопримечательности Мадейры: гора Кабо-Жирао, ботанический сад и сад орхидей и вулканические пещеры Сао-Винсенте. Кроме прочего, здесь стоит посетить крупный водноспортивный центр. В Фуншале стоит увидеть собор Се, монастырь Св. Клары и дворец-крепость Св. Лоренсо, посетить колоритный рынок Лаврадорес и погулять по паркам и садам. Must-see Порту-Санту: дом Колумба, церковь Ларго-до-Пелориньо, Ратуша, ветряные мельницы и ботсад Кинта-даш-Пальмейраш.
Кроме прочего, здесь стоит посетить крупный водноспортивный центр. В Фуншале стоит увидеть собор Се, монастырь Св. Клары и дворец-крепость Св. Лоренсо, посетить колоритный рынок Лаврадорес и погулять по паркам и садам. Must-see Порту-Санту: дом Колумба, церковь Ларго-до-Пелориньо, Ратуша, ветряные мельницы и ботсад Кинта-даш-Пальмейраш.
FUN&SUN | Отдых в Мадейре, Португалия : цены с перелетом из Москвы 2022-2023
Природа Мадейры уникальна- это заповедник, две трети территории которого объявлены охраняемой зоной и в которой встречается намало редких видов растений. Поклонники архитектуры так же не будут разочарованы: столицу острова, город Фуншал, не без оснований называют «Лиссабоном в миниатюре»; а в целом более чем 500-летнее культурно-историческое наследие острова отражено в многочисленных памятниках архитектуры. Уютный островок архипелага, Порту- Санту, расположенный к северо-востоку от Мадейры, славится своим изумительным по красоте девятикилометровым пляжем, еще Колумб пришел к выводу, что это идеальное место, чтобы забыть о повседневных проблемах и наслаждаться жизнью.
Расположение
Мадейра расположена в Атлантическом океане, приблизительно в 500 км к западу от Африканского побережья и в 1000 км к югу от Португалии, в 1,5 часах перелета из Лиссабона. Мадейра занимает площадь 741 кв. км. Архипелаг включает острова Мадейра, Порту-Санту, а так же необитаемые острова Дезерташ и Селваженш. Из 220.000 жителей архипелага проживают в столице -г. Фуншал.
Экскурсии
Мадейра — идеальное место для совершения пеших прогулок, а так же здесь открываются широчайшие возможности для занятия различными видами спорта- от параплана, скалолазания, горного велосипеда до верховой езды и тенниса. Так же здесь есть прекрасные поля для гольфа. В последнее время Мадейра становится первоклассным международным центром подводной рыбалки, где нередко вылавливают синих марлиней весом до 500 кг. И, конечно же, на Мадейре не будет скучно любителям серфинга, винд-серфинга и подводного плавания Прекрасным местом для занятий водными видами спорта является и Порту-Санту.
Кухня
Впечатляющее изобилие тропических фруктов, выращиваемых на богатых вулканических почвах, которые практически не требуют орошения химическими удобрениями, можно увидеть и попробовать только здесь! Так же на Ваш выбор широкий ассортимент свежей рыбы и даров моря. И конечно же, только здесь, Вам представится возможность отведать знаменитые вина, известные под общим названием «мадейра». Сухое Серсиал — прекрасный аперитив; полусухое Верделью; полусладкое Буал — отлично сочетается с сыром; сладкое Малвазия — незаменимо на десерт.
Климат
Уникальные климатические условия делают Мадейру идеальным местом для отдыха на природе в любое время года. Зимой здесь тепло, а летом никогда не бывает сильной жары. Так, средняя температура воздуха составляет 23 С летом и 19 С зимой. Среднегодовая температура морской воды так же держится в пределах 22 С и 17 С. В целом, здешний климат прекрасно тонизирует и восстанавливает силы.Рождение укулеле: как появился на свет один из самых универсальных струнных инструментов в истории
Первые укулеле появились в середине 1880-х годов. И не на Гавайях. Укулеле — инструмент, родом с острова — это правда, однако это не Гавайи, и даже не соседние острова в Тихом океане. Мадейра — небольшой гористый архипелаг в Атлантическом океане, принадлежащий Португалии — является настоящей родиной укулеле. Регион Мадейра обладает тропическим климатом и возник вследствие извержения вулкана. Мадейра славится своими густыми лесами (“Мадейра” в переводе с португальского означает “дерево”): раньше здесь была очень развита лесозаготовительная промышленность.
Два века назад Мадейра также была популярным местом среди туристов из Европы, которых привлекали шикарные виды и экзотическая флора. В самом крупном портовом городе Мадейры — Фуншале — местные музыканты частенько развлекали туристов своей игрой. Из-за жаркого климата в домах на архипелаге как правило не было оконных рам, поэтому музыку можно было слышать круглосуточно. Местные музыканты играли вальсы, мазурки и народные произведения на испанских гитарах и небольших четырёхструнных инструментах, которые назывались машете.
К сожалению, к середине 1800-х годов Мадейра перестала быть привлекательным местом для жизни: бедность, голод и серия природных катастроф, которая привела к коллапсу винного производства сделала архипелаг тем местом, с которого стремились уехать. Сотни безработных жителей искали возможность покинуть родину и начать новую жизнь в другой части мира. Интересно, что в то время, как жизнь на Мадейре угасала, она расцветала на другом острове в тысячах километров от неё: сахарное производство на Гавайях стремительно набирало обороты.
Интересно, что в то время, как жизнь на Мадейре угасала, она расцветала на другом острове в тысячах километров от неё: сахарное производство на Гавайях стремительно набирало обороты.
В 1874-м году гавайские плантаторы отправили на материк 25 тонн сахара. Существовала однако проблема: из-за европейской колонизации и привезенных на остров болезней местное население существенно сократилось, поэтому на острове не хватало людей, чтобы работать на плантациях. В отчаянии плантаторы начали искать рабочих по всему миру, и в конце концов новость об этом дошла и до Португалии. На Мадейре не составило труда найти мужчин и женщин, которые были готовы подписать трёхлетний контракт и отправиться работать на Гавайи. Помимо жалования в 10 долларов в месяц иммигранты получали жилье и питание, а также место на корабле, который отвёз бы их на новый остров.
Среди двадцати пяти тысяч жителей Мадейры, которые переселились на Гавайи в конце 1800-х было три плотника из Фуншала: сорокалетний Мануэль Нуньес, тридцатисемилетний Аугусто Диас и двадцативосьмилетний Жозе до Эспирито Санто. Вместе со своими семьями они собрали парусное судно и отправились в четырехмесячное путешествие к гавайскому острову Оаху. Никто из них тогда и не догадывался, что это приключение принесет им не только достаток, но и приведёт к появлению нового музыкального инструмента.
Бедные, утомлённые морем иммигранты в конце концов причалили в порту Гонолулу в тихую субботу августа 1879-го года. Как только они пришвартовались, один из пассажиров — музыкант Жоао Фернандес — принялся играть весёлую мелодию, отмечая успешное окончание их путешествия. Фернандес был талантливым музыкантом, который мог сыграть любую мелодию, услышав её лишь однажды, он играл на машете, заимствованной у одного из пассажиров. Очевидно, он был не единственным на острове музыкантом, который играл на машете: спустя несколько недель после прибытия переселенцев на остров, в местной газете вышла статья: “…переселенцы с Мадейры недавно приехали на наш остров и радуют жителей ночными уличными концертами. Музыканты отлично играют на своих странных инструментах, которые представляют собой некую смесь гитары и банджо, однако звучат очень приятно”.
Музыканты отлично играют на своих странных инструментах, которые представляют собой некую смесь гитары и банджо, однако звучат очень приятно”.
Нуньес, Диас и Санто попали работать на сахарные плантации на разные острова — Гавайи, Кауаи и Мауи. После того, как их контрактные обязательства были исполнены, все трое устремились в Гонолулу — столицу архипелага и центр торговли, намереваясь найти работу по их изначальной профессии. К счастью, в Гонолулу в то время развивалась мебельная промышленность, и в городе существовала около десятка различных мастерских. Нуньес и Санто получили работу в самом крупном мебельном магазине Гавайев — Pioneer Furniture House. Диас открыл свою небольшую мастерскую в китайском квартале города в 1884-м году: он делал не только мебель, но и музыкальные инструменты.
Спустя год Нуньес также открыл свой собственный магазин, всего в трёх домах от мастерской Диаса. Оба плотника рекламировали свои услуги через местную газету. Диас описывал себя как “мастера, который делает гитары, машете и прочие струнные инструменты”, Нуньес позиционировал себя в качестве “краснодеревщика, который делает всевозможные струнные инструменты, в частности — гитары и машете”. Вскоре и Санто открыл свой магазин в паре дверей от магазина Нуньеса. Помимо производства собственных инструментов все трое зарабатывали на жизнь перепродажей инструментов для масс-маркета, ремонтом, продажей струн и так далее. Диас даже давал уроки музыки.
Диас описывал себя как “мастера, который делает гитары, машете и прочие струнные инструменты”, Нуньес позиционировал себя в качестве “краснодеревщика, который делает всевозможные струнные инструменты, в частности — гитары и машете”. Вскоре и Санто открыл свой магазин в паре дверей от магазина Нуньеса. Помимо производства собственных инструментов все трое зарабатывали на жизнь перепродажей инструментов для масс-маркета, ремонтом, продажей струн и так далее. Диас даже давал уроки музыки.
Как же три этих простых плотника с Мадейры внезапно стали лютье? До сих пор неясно, делали ли кто-либо из них музыкальные инструменты до того, как они приехали на Гавайи: существует версия (однако, без прямых доказательств), что Нуньес мог быть родственником Октавиана Жоао Нуньеса да Пайксао (1812–1874) — одного из самых знаменитых лютье Мадейры. Более вероятная версия — Нуньес, Диас и Санто начали делать музыкальные инструменты в качестве стороннего бизнеса для заработка, продолжая заниматься своей основной профессией плотника. Несмотря на отсутствие должного образования в качестве лютье, по качеству их инструментов можно было сказать, что они точно знали, что делали.
Несмотря на отсутствие должного образования в качестве лютье, по качеству их инструментов можно было сказать, что они точно знали, что делали.
Первое упоминание укулеле как отдельного инструменте встречается в газете спустя около десяти лет после прибытия на остров переселенцев с Мадейры. Кто же сделал первое укулеле? Честно говоря, до конца неясно. Все три мастера делали машете, которые во многом походили на укулеле, а Санто утверждал в своей рекламе, что “может сделать гитару любого размера”. Нуньес заявлял, что именно он изобрёл укулеле, делая на этом акцент в своей рекламе.
Какую бы роль не сыграли в появлении на свет укулеле Нуньес, Диас и Санто, скорее всего первая укулеле представляла собой собой гибридный инструмент: сочетание машете и другого небольшого португальского струнного инструмента — пятиструнного ражанью. За основу укулеле была взята форма корпуса машете и его небольшой размер, а также 17-ладовый гриф. Строй машете (D-G-B-D) однако отличался от строя укулеле: укулеле настраивали в строй ражанью (без пятой струны) — G-C-E-A.
Почему был выбран строй G-C-E-A?
“Когда и почему укулеле получила отличный от машете строй — один из тех вопросов, который до сих пор остаётся без ответа”, — пишет историк музыки Джон Кинг в своей книге “ The Ukulele: A History”. Ещё одним важным отличием гавайской укулеле от его португальских родственников стал выбор пород древесины для их производства. Машете и ражанью чаще всего изготавливали из ели (в качестве материала верхней деки) и можжевельника (в качестве материала корпуса) или других лёгких пород. Практически все ранние укулеле были сделаны целиком из коа: древесины золотисто-коричневого цвета, почитаемой жителями Гавайев, которая традиционно использовалась для производства дорогой мебели. Санто выбирал для своих укулеле первоклассную фигурную коа и использовал элементы декора, которые часто встречались на машете. Корпусы его укулеле были невероятно тонкими, что делало инструменты особенно лёгкими, однако они звучали удивительно громко и богато для своего небольшого размера.
То, что укулеле получило своё особенное название — вещь, которая помогает отличить информацию о ранних укулеле от информации о прочих похожих инструментах, однако как и почему укулеле было названо “укулеле” — ещё одна загадка. Существует несколько теория, но одна из них — самая разумная: слово “укулеле” в гавайском языке существовало и до появления нового инструмента. В словаре 1865-го года встречается слово “укулеле”, его значение — “кошачья блоха” — вредное насекомое, которое было завезено на остров десятилетия назад. Около 1900-го года американский писатель Джек Лондон писал, что “укулеле — это “прыгающая блоха” по-гавайски, однако это также и музыкальный инструмент, похожий на гитару”.
Какой бы ни была этимология слова укулеле на самом деле, новый инструмент быстро получил популярность благодаря, отчасти, высшим чинам островов: Дэвид Калакауа, последний король Гавайев, его жена — королева Эмма и будущая королева — Лилиуокалани (которая сочинила “Aloha Oe“ — священную для жителей Гавайев песню) — все они были музыкантами и сторонниками искусства. Их поддержка и продвижение укулеле помогли остальным местным жителям взять инструмент в руки и создавать свою музыку и стили игры. Помимо того, что Калакауа приглашал укулелистов играть на королевских приёмах, он научился играть на укулеле самостоятельно и часто выступал на встречах со своими друзьями.
Их поддержка и продвижение укулеле помогли остальным местным жителям взять инструмент в руки и создавать свою музыку и стили игры. Помимо того, что Калакауа приглашал укулелистов играть на королевских приёмах, он научился играть на укулеле самостоятельно и часто выступал на встречах со своими друзьями.
“Мы приходили в гости в бунгало короля, — вспоминает музыкант Жоао Фернандес в интервью журналу Paradise of the Pacific в 1922-м году. — Много народу приходило. Было много местных, много музыки, много еды, много напитков. И король Калакауа играл для нас!”.
Очевидно именно так король Дэвид получил прозвище, которое до сих пор используется на Гавайях — “Весёлый Монарх”. По мере того, как на Гавайях появлялось новое поколение лютье — включая Джонаса Кумулае, который сделает укулеле известным по всему миру, выставив его на выставке Pan Pacific International Exhibition в Сан Франциско в 1915-м году — предыдущее поколение постепенно отходило от дел. К 1900-му году Санто закрыл свою мастерскую, но продолжал работать дома ещё несколько лет, прежде чем умер. Диас потерял свою мастерскую в безжалостном пожаре, который уничтожил большую часть китайского квартала примерно в то же время. Нуньес, самый продуктивный лютье из них троих, продолжил изготавливать инструменты ещё долгое время. Он передал свои знания и опыт большому количеству молодых лютье, включая своего сына Леонардо, который возглавлял фабрику Нуньесов в Лос Анджелесе до 1930-го года. Другой ученик Мануэля — Самуэль Камака открыл свою мастерскую в 1916-м году. Теперь, практически 100 лет спустя, мастерская Kamaka Ukulele and Guitar Works в Гонолулу олицетворяет собой наследие трёх португальских иммигрантов, которые навсегда изменили гавайскую музыку, подарив острову “прыгающую блоху”.
К 1900-му году Санто закрыл свою мастерскую, но продолжал работать дома ещё несколько лет, прежде чем умер. Диас потерял свою мастерскую в безжалостном пожаре, который уничтожил большую часть китайского квартала примерно в то же время. Нуньес, самый продуктивный лютье из них троих, продолжил изготавливать инструменты ещё долгое время. Он передал свои знания и опыт большому количеству молодых лютье, включая своего сына Леонардо, который возглавлял фабрику Нуньесов в Лос Анджелесе до 1930-го года. Другой ученик Мануэля — Самуэль Камака открыл свою мастерскую в 1916-м году. Теперь, практически 100 лет спустя, мастерская Kamaka Ukulele and Guitar Works в Гонолулу олицетворяет собой наследие трёх португальских иммигрантов, которые навсегда изменили гавайскую музыку, подарив острову “прыгающую блоху”.
Укулеле Kala разработаны в лучших традициях легендарных гавайских инструментов, сочетая в себе, тем не менее, современные элементы для того, чтобы вам было максимально комфортно играть на вашем инструменте.
Цветы архипелага Мадейра | Выпуск 37/3 2010
По следам Колумба, ради открытия архипелага редкой красоты, с пышными лесами и богатыми местными традициями.Татьяна Песчанская, врач, кандидат медицинских наук, страстный путешественник и наш постоянный автор
Архипелаг Мадейра состоит из двух главных островов: Мадейры, благодаря которому он получил свое название, и Порту-Санту, первому острову, открытому португальцами во время исследовательских и колонизаторских морских путешествий, начавшихся в XV веке. Столица архипелага – Фуншал, город, основанный в 1425 году в красивой бухте, окруженной естественным амфитеатром гор. Согласно хроникам той эпохи, первый город построен «в прекрасной долине, где в изобилии растут необычные деревья и фенхель, спускающиеся до самого моря», как писал Гаспар Фрутуозу.
Мадейра, длиной 57 км и шириной 22 км, с населением 260 тысяч человек – это политико-административное образование, созданное рамках Конституции Португальской республики. Сегодня, вследствие модернизации и строительства новых дорог и гостиниц, Мадейра и Порту-Санту утвердились как первоклассные туристические объекты, которые привлекают отдыхающих со всех концов света и во все времена года. В самом деле, благодаря глубоким долинам, пышно разросшимся лесам и цветам на Мадейре можно полюбоваться потрясающими пейзажами, поистине уникальным своеобразием природы и, к тому же, насладиться великолепным климатом, тихой и успокаивающей атмосферой.
Сегодня, вследствие модернизации и строительства новых дорог и гостиниц, Мадейра и Порту-Санту утвердились как первоклассные туристические объекты, которые привлекают отдыхающих со всех концов света и во все времена года. В самом деле, благодаря глубоким долинам, пышно разросшимся лесам и цветам на Мадейре можно полюбоваться потрясающими пейзажами, поистине уникальным своеобразием природы и, к тому же, насладиться великолепным климатом, тихой и успокаивающей атмосферой.
Леса на скалах
Рассказ о Мадейре означает разговор о её городах, базальтовых скалах и галечных пляжах, также базальтового происхождения. Остров покрыт своеобразным «туземным» лесом, называемым Лауриссильва («Лавровый лес»). Подобные леса, распространенные прежде и на европейском континенте, стали жертвой последующих оледенений и выжили только в регионе Макронезии (по-гречески – «блаженные острова»), включающем Мадейру, Ахоры, Канары и КабоВерде (острова Зеленого Мыса).
Именно на Мадейре эта почти ископаемая, но живая растительность занимает 22 тысячи гектаров. Леса растут на высоте от 300 до 1300 метров над уровнем океана и играют главную роль в защите почвы, способствуя сохранению дождевой воды. Лес Лауриссильва целиком входит в состав Национального Парка Мадейры со статусом «строгого заповедника» и тщательно оберегается.
Леса растут на высоте от 300 до 1300 метров над уровнем океана и играют главную роль в защите почвы, способствуя сохранению дождевой воды. Лес Лауриссильва целиком входит в состав Национального Парка Мадейры со статусом «строгого заповедника» и тщательно оберегается.
Благодаря своему субтропическому климату Мадейра представляет собой идеальное место для выращивания цветов. Цветы стали визитной карточкой острова. Прославленные незабываемыми запахами ночи на Мадейре воспеты поэтами и композиторами. Остров знаменит многочисленными парками, наполненными удивительным разнообразием растений и красочных цветов на радость посетителям, которые ценят не только романтическое очарование, но и научную ценность местной флоры. Недаром на остров, получивший название «Сад на берегу моря», приезжают из разных стран научные группы, включающие специалистов из таких областей, как ботаника, садоводство и цветоводство.
В прямом смысле слова цветущий город Фуншал в последние годы получил множество разнообразных европейских премий, что только повысило его привлекательность.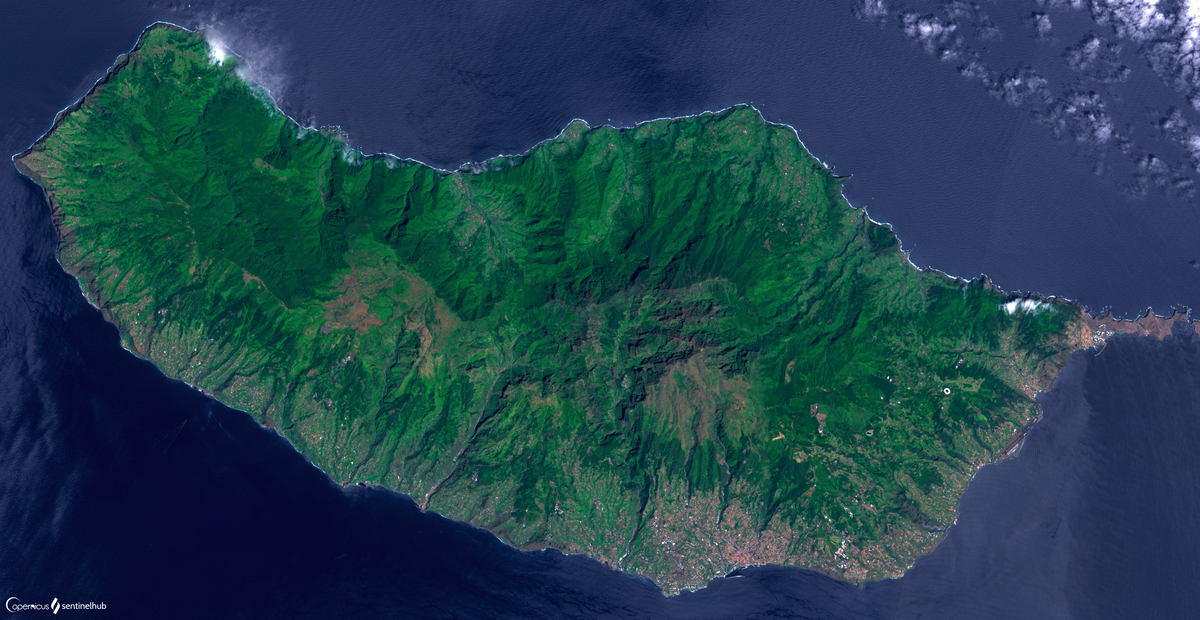 Среди самых впечатляющих нельзя не заметить «райских цветов», великолепного и очень выносливого растения, типичного для тропического острова, произрастающего вместе с антуриумом и протеей (эндемический вид на Мадейре и в Южной Африке). Всё это, наряду с огромным числом диких видов, которые усеивают весь остров, меняет его внешний вид в соответствии с временами года. Цветы на Мадейре – это часть повседневной жизни, прекрасно выражающая гостеприимную природу и радующая остров и его обитателей.
Среди самых впечатляющих нельзя не заметить «райских цветов», великолепного и очень выносливого растения, типичного для тропического острова, произрастающего вместе с антуриумом и протеей (эндемический вид на Мадейре и в Южной Африке). Всё это, наряду с огромным числом диких видов, которые усеивают весь остров, меняет его внешний вид в соответствии с временами года. Цветы на Мадейре – это часть повседневной жизни, прекрасно выражающая гостеприимную природу и радующая остров и его обитателей.
Щедрая лоза
Из всех продуктов, производимых на архипелаге, вина Мадейры пользуются воистину мировой известностью. По свидетельствам историков, виноделие возникло на острове в эпоху португальской колонизации. Первыми, кто стал торговать мадерой, были иезуиты, владевшие обширными виноградниками: продажа этого крепкого бодрящего вина позволила монашескому ордену утвердить свое экономическое могущество и приобрести большое влияние в социальной сфере.
Посетивший Мадейру в 1455 году путешественник – венецианец Луиджи ди Казамосто, восторгался местным вином, особенно мальвазией критского происхождения. Прославленное вино в середине XVII века было воспето Уильямом Шекспиром устами своего Фальстафа. Вино, которое, в доказательство важности его экспорта во времена войны между кланами Алой и Белой Розы, стало главным «действующим лицом» этих печальных событий: король Эдвард II пошел против своего брата, герцога Кларенского, и тот решил покончить с собой, утопившись в бочке вина с Мадейры. Однако в начале XVIII века производство мадеры перешло в руки англичан, чьи корабли стали систематически причаливать на пути к Антильским островам. Полученное путем брожения свежего виноградного сока вино мадера – довольно сладкое, благодаря наличию в нем определенного количества остаточного сахара. Сейчас отборный виноград с заботливо выращенных виноградников давят механически (прежде сок выжимали ногами в прессах больших виноградарских хозяйств, с частными винокурнями). Затем сусло подвергается спиртовому брожению – это первый этап длительного процесса, который завершается очисткой в «баррикиш».
Прославленное вино в середине XVII века было воспето Уильямом Шекспиром устами своего Фальстафа. Вино, которое, в доказательство важности его экспорта во времена войны между кланами Алой и Белой Розы, стало главным «действующим лицом» этих печальных событий: король Эдвард II пошел против своего брата, герцога Кларенского, и тот решил покончить с собой, утопившись в бочке вина с Мадейры. Однако в начале XVIII века производство мадеры перешло в руки англичан, чьи корабли стали систематически причаливать на пути к Антильским островам. Полученное путем брожения свежего виноградного сока вино мадера – довольно сладкое, благодаря наличию в нем определенного количества остаточного сахара. Сейчас отборный виноград с заботливо выращенных виноградников давят механически (прежде сок выжимали ногами в прессах больших виноградарских хозяйств, с частными винокурнями). Затем сусло подвергается спиртовому брожению – это первый этап длительного процесса, который завершается очисткой в «баррикиш».
Вообще, все местные вина подвергаются процессу «иштуфажень» – традиционному на Мадейре винодельческому приему, заключающемуся в их нагреве и выдержке при постоянной температуре 50°С. После этого вино выдерживается в чанах не менее 90 дней. Этому процессу не подвергается ряд «резервов», в частности вина определенных урожаев, требующие медленного старения в бочках.
После этого вино выдерживается в чанах не менее 90 дней. Этому процессу не подвергается ряд «резервов», в частности вина определенных урожаев, требующие медленного старения в бочках.
В традиционных винокурнях можно отыскать старые вина исключительного качества, стоимость которых растет с возрастом. Их дегустация – настоящее удовольствие, и к ней можно приступить в любое время дня. Особенности вин Мадейры, так же как и манера их подавать, меняются в зависимости от виноградной лозы, из которой они изготовлены. Изысканное вино «Мадера», и поныне изготавливаемое строго по традиционным рецептам, ценимое повсюду и доставляемое во все уголки планеты, по справедливости пользуется громкой славой в мире.
Ботанический сад
Уникальным по своей природе является Ботанический сад, разбитый на Мадейре в 1960 году. Засаженная площадь сада составляет 35 тысяч квадратных метров, представляя собою удивительное зрелище с гармоничными формами и целой палитрой красок.
Отсюда открывается прекрасная панорама на бухту и порт. Здесь собрано более двух тысяч видов экзотических растений со всех континентов, в том числе – орхидеи, антуриумы, райские цветы, кактусы, пальмы, папоротники. Именно это изобилие видов вместе с множеством самых прекрасных цветов острова и придает Ботаническому саду поистине уникальную красоту. По соседству с Ботаническим садом находится орнитологический заповедник, где бесчисленные экзотические птицы живут практически на свободе, вместе с множеством видов порхающих повсюду мелких птах. В парке Лойру, рядом с Ботаническим садом, можно полюбоваться попугаями-ара, какаду и многими другими экзотическими видами.
Здесь собрано более двух тысяч видов экзотических растений со всех континентов, в том числе – орхидеи, антуриумы, райские цветы, кактусы, пальмы, папоротники. Именно это изобилие видов вместе с множеством самых прекрасных цветов острова и придает Ботаническому саду поистине уникальную красоту. По соседству с Ботаническим садом находится орнитологический заповедник, где бесчисленные экзотические птицы живут практически на свободе, вместе с множеством видов порхающих повсюду мелких птах. В парке Лойру, рядом с Ботаническим садом, можно полюбоваться попугаями-ара, какаду и многими другими экзотическими видами.
Пейзажи Монти и августейшие особы
Одно из самых живописных местечек острова – Монти (Monte). Именно здесь, на заре колонизации, сходились жители острова с тем, чтобы помолиться в часовне, посвященной Божьей Матери Воплощения. Монти, стоящий всего в нескольких километрах от главного порта, расположен в изумительной природной среде на солнечных склонах гор, окружавших широким амфитеатром Фуншал.
Чарующий вид, особенно если смотреть со стороны моря, на протяжении столетий прославляли художники, причислив его к прекраснейшим пейзажам мира. Монти – это селение, полное фруктовых садов, которые перемежаются с полями, принадлежавшими старинным дворянским семействам. В XIX веке Монти как место летнего отдыха особенно ценили благородные семейства Мадейры, построившие здесь уютные виллы. В дальнейшем отдых на Монти привлек и многочисленных иностранцев, в частности, британцев и немцев, а также тех купцов, что останавливались здесь на пути к Антильским островам и в Африку.
В Монти выстроены первые на острове отели – на участках, обсаженных деревьями в североевропейском стиле. В XIX веке остров и его столица стали местом романтического отдыха, прежде всего благодаря визитам многочисленных коронованных особ: австрийской императрицы Елизаветы (знаменитой Сисси), английской королевы Аделаиды, бельгийской принцессы Шарлотты и многих других королевских домов Европы. Еще большую известность остров приобрел тогда, когда наряду с пляжными и бальнеологическими учреждениями, его достопримечательности стали неотъемлемой частью маршрута «Гран тур» (образовательного путешествия юных европейских джентльменов). Интерес туристов, посетивших остров Мадейра, вызывали рассказы о пребывании императрицы Сисси на Мадейре (австрийская императрица Елизавета). Хотя пребывание императрицы Сисси на Мадейре хорошо отражено в документах того времени, многое все еще покрыто тайной.
Интерес туристов, посетивших остров Мадейра, вызывали рассказы о пребывании императрицы Сисси на Мадейре (австрийская императрица Елизавета). Хотя пребывание императрицы Сисси на Мадейре хорошо отражено в документах того времени, многое все еще покрыто тайной.
Елизавету (Элизабет) Виттельсбах, императрицу Австрии, считали самой красивой государыней мира: при росте 172 см она весила около 50 кг, а окружность её талии не превышала 50 см. Сисси много занималась гимнастикой в специально оборудованном для этого зале и часто ездила верхом (её называли лучшей наездницей того времени). В 1859 году она решила оставить мужа и сыновей, и уехала в Германию, в Поссенхофен, откуда стала выезжать в длительные и частые путешествия. В итоге, государыня добралась до Мадейры, где жила почти в затворничестве, прежде всего потому, что лечилась от туберкулеза. Действительно, благодаря выгодному географическому положению и здоровому климату, остров Мадейра и сегодня является лучшим местом для лечения, Что, кстати, знали европейские придворные и члены самых состоятельных семейств (их на Мадейре было немало).
Когда в 1861 году императрица проживала на острове, по распоряжению короля Португалии ей было предоставлено крыло во дворце Санлореску. Однако императрица предпочла Кинита Вижия – романтического вида особняк с широкими верандами и большим садом с видом на море. Здесь сейчас находятся отель и казино.
На Мадейре погребен последний австрийский император Карл I. Утром 19 ноября 1921 года на борту британского крейсера «Кардифф» на Мадейру прибыл бывший император Австро-Венгрии Карл I Габсбург. Семья Карла воссоединилась и обосновалась в Монти, в Доме банкира Луиса да Роша Машаду. Там 1 апреля 1922 года после тяжелой болезни и скончался последний император Австрии. Его похоронили через 4 дня на кладбище у приходской церкви в Монти. После этого печального события экс-императрица с детьми решила навсегда покинуть остров. Особое впечатление оставляет посещение Кафедрального собора Фуншала (1514 год) и музея сакрального искусства. В Епархиальном музее сакрального искусства хранится собрание большой художественной ценности. Здесь есть великолепные коллекции произведений фламандских живописцев XVI – XVIII веков.
Здесь есть великолепные коллекции произведений фламандских живописцев XVI – XVIII веков.
Присутствие на архипелаге фламандского искусства весьма знаменательно: шедеврами этой прославленной школы (живопись, скульптура, изделия из серебра, ювелирные украшения) можно полюбоваться в разных местах острова, но прежде всего, в залах музея. Здесь также выставлена живопись по дереву, созданная великими художниками того времени, триптихи и заалтарные украшения, где сочетаются живопись и скульптура. В музее собраны ювелирные изделия для литургических облачений и картины португальских художников XV – XVIII веков. Здесь также хранится часть сокровищницы Собора, в том числе такие уникальные произведения, как золотой крест для процессий, подаренный королем Мануэлом I и предметы из серебра – металла, очень распространенного в XVI – XVII веках, благодаря широкой торговле кастильских «серебряных» галлонов.
«Северный сад» – Сантана
Неотъемлемым этапом для всех, путешествующих по Мадейре, является посещение Сантаны. Красота её цветущих равнин, богатство флоры и разнообразие фауны – все это складывается в изумительную фреску, исполненную мира и спокойствия и передающую особенное очарование этой части острова. Сантану окрестили «северным садом». Вполне заслуженное имя, потому что здесь во всех домах есть сады. Называемые «кантейрум» (клумба или газон), изобилием цветов и ароматов они очаровывают путешественников в любое время года. Их облик говорит об увлеченности и артистизме жителей этих мест, которые повсюду разбивают сады. Ливадаш – ирригационные каналы, прорытые в горах, – еще одна особенность пейзажа Мадейры. Сегодня они дают возможности любителям природы совершить увлекательные прогулки в глубину туземного леса, увидеть живописные пейзажи, где вдоль тропинок в изобилии растут волчья ягода, кизил, гортензия, самшит.
Красота её цветущих равнин, богатство флоры и разнообразие фауны – все это складывается в изумительную фреску, исполненную мира и спокойствия и передающую особенное очарование этой части острова. Сантану окрестили «северным садом». Вполне заслуженное имя, потому что здесь во всех домах есть сады. Называемые «кантейрум» (клумба или газон), изобилием цветов и ароматов они очаровывают путешественников в любое время года. Их облик говорит об увлеченности и артистизме жителей этих мест, которые повсюду разбивают сады. Ливадаш – ирригационные каналы, прорытые в горах, – еще одна особенность пейзажа Мадейры. Сегодня они дают возможности любителям природы совершить увлекательные прогулки в глубину туземного леса, увидеть живописные пейзажи, где вдоль тропинок в изобилии растут волчья ягода, кизил, гортензия, самшит.
Пробравшись в самый центр Сантаны, можно открыть для себя дикую красоту края Ашада-ду – Тейшера, нередко окутанного густым туманом! После нескольких минут ходьбы по аккуратно проложенной тропинке – вот она, величественная вершина Пику-Руйву, самая высокая на острове, подлинный шедевр природы и непреложная цель гостей Мадейры. Можно остановиться в маленькой гостиницу у самого Пику Руйву, чтобы утром увидеть незабываемое зрелище восхода солнца над облаками на отметке в 2 тысячи метров.
Можно остановиться в маленькой гостиницу у самого Пику Руйву, чтобы утром увидеть незабываемое зрелище восхода солнца над облаками на отметке в 2 тысячи метров.
О веселом нраве, танцах и вышивках островитян
Все на Мадейре повествует о древних традициях, даже местная экономика, с её историческими ресурсами – сельским хозяйством, разведением скота с его древними обрядами и, конечно, рыбной ловлей, с типичными лодками, бороздящими океанские воды с незапамятных времен. На Мадейре ремесленные изделия становятся истинными шедеврами, известными во всем мире. Достаточно упомянуть необыкновенные плетеные вещи, изготовленные из местных растений с потрясающей ловкостью, и превосходные вышивки, настоящая специализация женщин с Мадейры – это не только два столпа местной экономики, но и составляющая часть богатейшего островного фольклора.
На Мадейре вышивают исключительно вручную. И в Европе, и в Америке вышивка с Мадейры считалась самой красивой в мире. Её высоко ценили эксперты и часто заказывали королевские семейства. В середине XIX века англичанка мисс Фелпс впервые познакомила с этими вышивками викторианскую Англию. Эти творения, вышитые крестьянками острова в эпоху, когда сельское хозяйство было единственным средством существования, очаровали придворных дам и представительниц высшего общества: благодаря своей рафинированности и тщательной проработке деталей вышивки с Мадейры стали пользоваться успехом.
В середине XIX века англичанка мисс Фелпс впервые познакомила с этими вышивками викторианскую Англию. Эти творения, вышитые крестьянками острова в эпоху, когда сельское хозяйство было единственным средством существования, очаровали придворных дам и представительниц высшего общества: благодаря своей рафинированности и тщательной проработке деталей вышивки с Мадейры стали пользоваться успехом.
Для фольклора Мадейры характерны веселая музыка и живописные выразительные танцы. Если отдельные песни, явно мавританского происхождения, навевают легкую меланхолию, то инструментальная музыка, оживляющая традиционные праздники и фольклорные фестивали, выражает обычно праздничное настроение радость жизни. Самый известный танец на Мадейре – это, несомненно, балинько.
Есть и другой, более народный и более крестьянский танец – бринку, под ритм которого во время традиционных праздников соревнуются сельское население, выполняя самые причудливые танцевальные па, сопровождающиеся пением. Жители Мадейры отличаются открытостью и сердечностью, с туристами ведут себя приветливо и дружелюбно. Впрочем, и тепло, и гостеприимство теперь известны в любой части мира: в силу того, что в основе экономики острова лежат, прежде всего, туризм и торговля, на острове понимают, что должны хорошо принимать гостей.
Жители Мадейры отличаются открытостью и сердечностью, с туристами ведут себя приветливо и дружелюбно. Впрочем, и тепло, и гостеприимство теперь известны в любой части мира: в силу того, что в основе экономики острова лежат, прежде всего, туризм и торговля, на острове понимают, что должны хорошо принимать гостей.
Перед гостями острова предстает первоклассная инфраструктура гостиниц, отличная организованность – вот секреты недавнего и быстрого развития туризма, чему способствовало устройство превосходных полей для гольфа. Два поля для гольфа расположены в нескольких минутах езды от Фуншала и от Канису, где находятся лучшие гостиничные комплексы острова. Площадка для гольфа в Санту да Серра самая важная для Мадейры, её разметил Роберт Гирент Джонс. Здесь ежегодно проходит весенний турнир Madera Open, входящий в систему P.Y.A/ European Tour. Площадка в Пальмейру расположена среди гор, возвышающихся над Фуншалом, откуда открывается чарующий вид на бухту и город. Её разметил Кейбилл Робинсон. Она находится в огромном парке с пышной растительностью, откуда открывается великолепный вид. В огромном парке здесь можно наблюдать поразительное разнообразие экзотических птиц (орнитологи насчитывают 291 вид). На этих площадках пробовали свои силы крупнейшие спортсмены мира.
Она находится в огромном парке с пышной растительностью, откуда открывается великолепный вид. В огромном парке здесь можно наблюдать поразительное разнообразие экзотических птиц (орнитологи насчитывают 291 вид). На этих площадках пробовали свои силы крупнейшие спортсмены мира.
Очень интересным и захватывающим впечатлением было посещение Mercado dos Zavradores (Рынок тружеников). На весьма оживленном по утрам базаре с рядами прилавков можно увидеть сцены, полные жизни, красок, запахов, а также потоки покупателей. Это место относится к самым привлекательным для приезжающих на Мадейру: они могут здесь буквально прикоснуться руками к бесконечному разнообразию продуктов острова.
Живописные прилавки, где предлагаются любые разновидности свежих продуктов, представляют собой поистине захватывающую картину. На Рыбном рынке на гранитных скамьях разложены настоящие богатства моря. Здесь продаются все виды свежей рыбы, в особенности тунец и «эшпада» (знаменитая плоская черная рыба), самые распространенные на Мадейре, а также моллюски и ракообразные, которых рыбаки привозят из богатых вод вокруг острова Мадейра или с Азорских островов. Всё на Мадейре повествует о древних традициях, даже местная экономика, с её историческими ресурсами – сельским хозяйством, разведением скота с его древними обрядами и, конечно, рыбной ловлей, с типичными лодками, бороздящими океанские воды с незапамятных времен.
Всё на Мадейре повествует о древних традициях, даже местная экономика, с её историческими ресурсами – сельским хозяйством, разведением скота с его древними обрядами и, конечно, рыбной ловлей, с типичными лодками, бороздящими океанские воды с незапамятных времен.
Традиционной культурой на Мадейре является выращивание бананов, манго и папайи, что дает средства к существованию многим семьям. По субботам и воскресеньям крестьяне собираются на большой площади города, чтобы продать плоды своих трудов: зелень, овощи, фрукты, цветы и т.д. Здесь устанавливаются столы, где можно пить и есть, как на всех португальских праздниках. Здесь можно отведать типичные блюда островной кухни, традиционные кебабы с лавровым листом и сладости. Это идеальное место для покупок, разнообразие и качество которых прославили Мадейру.
Прозрачное и чистое море с волнами, доставляющими радость поклонникам виндсерфинга, нетронутая природа, бескрайние густые леса, зеленые луга с разноцветными клумбами – всё это на архипелаге, по достоинству признанном райским уголком, где самая высокая продолжительность жизни в мире. И еще – остров Мадейра по праву считается островом вечной весны и молодости. Приехав сюда, хочется остаться навечно, соединившись с природой, и наслаждаться жизнью.
И еще – остров Мадейра по праву считается островом вечной весны и молодости. Приехав сюда, хочется остаться навечно, соединившись с природой, и наслаждаться жизнью.
Мадейра | Описание и факты
Острова Мадейра , Португалия Arquipélago da Madeira , архипелаг вулканического происхождения в северной части Атлантического океана, принадлежащий Португалии. В его состав входят два обитаемых острова, Мадейра и Порту-Санту, и две необитаемые группы, Дезертас и Сельвагенс. Острова — это вершины гор, основания которых лежат на бездонном дне океана. В административном отношении они образуют автономный регион Мадейра. Столица региона Фуншал расположена на острове Мадейра.
Узнайте об истории и природных ландшафтах острова Мадейра
Обзор острова Мадейра, Португалия.
Contunico © ZDF Enterprises GmbH, Майнц Посмотреть все видео к этой статье Остров Мадейра, самый большой из группы, имеет длину 34 мили (55 км), максимальную ширину 14 миль (22 км) и береговую линию около 90 миль (144 км) и возвышается в центре до пика Руйво (6 106 футов [1861 метр] над уровнем моря). Большая часть внутренних районов на высоте более 3000 футов (900 метров) необитаема и необрабатываема; общины разбросанных хижин строятся обычно либо в устьях оврагов, либо на склонах, спускающихся с гор к побережью.
Большая часть внутренних районов на высоте более 3000 футов (900 метров) необитаема и необрабатываема; общины разбросанных хижин строятся обычно либо в устьях оврагов, либо на склонах, спускающихся с гор к побережью.
Британская викторина
Острова и архипелаги
Из чего состоят Мальдивские острова? Какой самый большой в мире архипелаг? Разберитесь в фактах об островах по всему миру.
Остров Порту-Санту находится примерно в 26 милях (42 км) к северо-востоку от Мадейры. Его главный город, Вила-де-Порто-Санту, местные жители называют Вила.На каждом конце острова есть холмы, из которых пик Фачо, самый высокий, достигает 1696 футов (515 метров). Культуры включают пшеницу, виноград и ячмень.
Пустыня находится примерно в 11 милях (18 км) к юго-востоку от Мадейры и состоит из трех островков: Чао, Бужио и Дезерта-Гранди, а также Прего-ду-Мор у северной оконечности острова Чао. Кролики и дикие козы живут на скудных пастбищах и привлекают случайных охотников в когда-то населенную Дезерта-Гранде. Сельвагены, или Острова Спасения, представляют собой три необитаемых скалы, расположенные в 156 милях (251 км) к югу от Мадейры, между последней и Канарскими островами.Самый большой имеет окружность около 3 миль (5 км).
Кролики и дикие козы живут на скудных пастбищах и привлекают случайных охотников в когда-то населенную Дезерта-Гранде. Сельвагены, или Острова Спасения, представляют собой три необитаемых скалы, расположенные в 156 милях (251 км) к югу от Мадейры, между последней и Канарскими островами.Самый большой имеет окружность около 3 миль (5 км).
Узнайте о проблемах выращивания на террасе на острове Мадейра
Обзор выращивания на террасе с акцентом на острове Мадейра.
Contunico © ZDF Enterprises GmbH, Майнц Просмотреть все видео к этой статье Сельское хозяйство долгое время было доминирующим видом деятельности на островах Мадейра и Порту-Санту. Культуры, исторически выращиваемые на островах, включают сладкий картофель; тыквы разных видов; кало, или таро, завезенный с островов Тихого океана; большинство кулинарных овощей Европы; хлопья; сладкая палочка; и различные фрукты, такие как апельсины, лимоны, гуава, манго, мушмула, заварные яблоки, инжир, ананасы и бананы. В настоящее время преобладают банановые плантации и виноградники, а бананы и одноименное вино Мадейра входят в число наиболее важных статей экспорта. Другие виды экономической деятельности включают переработку сахара, рыболовство и ремесла, такие как деревообработка, плетение и вышивка, последнее из которых было представлено Мадейре в 1850-х годах Элизабет Фелпс, дочерью английского торговца вином. К началу 21 века туризм добился значительных успехов. Площадь 309 квадратных миль (801 кв. км). Поп. (2001) 245 011; (2011) 267 785; (оценка 2018 г.) 253 945.
В настоящее время преобладают банановые плантации и виноградники, а бананы и одноименное вино Мадейра входят в число наиболее важных статей экспорта. Другие виды экономической деятельности включают переработку сахара, рыболовство и ремесла, такие как деревообработка, плетение и вышивка, последнее из которых было представлено Мадейре в 1850-х годах Элизабет Фелпс, дочерью английского торговца вином. К началу 21 века туризм добился значительных успехов. Площадь 309 квадратных миль (801 кв. км). Поп. (2001) 245 011; (2011) 267 785; (оценка 2018 г.) 253 945.
Карта островов Мадейра ( c. 1900), из 10-го издания Британской энциклопедии .
Британская энциклопедия, Inc.Мадейра и Азорские острова | Encyclopedia.com
Архипелаг Мадейра, расположенный примерно в 350 милях от северо-западного побережья Африки и в 520 милях к юго-западу от самой Португалии, состоит из главного острова Мадейра и меньшего острова, Порту-Санту, в двадцати пяти милях к северо-востоку. Хотя ранее это было известно, острова были необитаемы, когда на них заявили права португальцы около 1419 года. Португальцы начали заселять их около 1425 года. Древесина, особенно кедр и тис, стали важным экспортным товаром, наряду с такими красителями, как драконья кровь, ятрышник и вайда. Богатая вулканическая почва стала еще более
плодородный, сжигая большую часть покрытого деревьями острова. Поскольку Мадейра была очень гористой, пришлось построить террасы. Пшеница стала важным более ранним продуктом. Подсчитано, что с 1450 по 1470 год Мадейра производила от 3000 до 3500 тонн в год.Были посажены виноградные лозы, завезен сахар, и примерно к 1452 году на Мадейре появился первый сахарный завод. Вскоре сахар стал основным продуктом архипелага и стал продаваться по всей Европе. Однако к середине шестнадцатого века сахарный бум начал заканчиваться, и вино постепенно вытеснило его в качестве основного экспорта острова.
Хотя ранее это было известно, острова были необитаемы, когда на них заявили права португальцы около 1419 года. Португальцы начали заселять их около 1425 года. Древесина, особенно кедр и тис, стали важным экспортным товаром, наряду с такими красителями, как драконья кровь, ятрышник и вайда. Богатая вулканическая почва стала еще более
плодородный, сжигая большую часть покрытого деревьями острова. Поскольку Мадейра была очень гористой, пришлось построить террасы. Пшеница стала важным более ранним продуктом. Подсчитано, что с 1450 по 1470 год Мадейра производила от 3000 до 3500 тонн в год.Были посажены виноградные лозы, завезен сахар, и примерно к 1452 году на Мадейре появился первый сахарный завод. Вскоре сахар стал основным продуктом архипелага и стал продаваться по всей Европе. Однако к середине шестнадцатого века сахарный бум начал заканчиваться, и вино постепенно вытеснило его в качестве основного экспорта острова.
Сахару, конечно же, требовалась рабочая сила. Сначала использовались гуанчи с Канарских островов и рабы-мусульмане из Северной Африки, а затем черные рабы после путешествий, спонсируемых принцем Генрихом (1394–1460). В то время как сахар был королем, рабская часть населения архипелага составляла примерно 10 процентов. К 1460-м годам было подсчитано, что на Мадейре проживало две тысячи жителей. К началу шестнадцатого века здесь проживало двадцать тысяч человек, в том числе около двух тысяч рабов. Тем временем, чтобы поощрить колонизацию, главный остров Мадейра был разделен на два наследственных владения лордов, Фуншал и Машику, с обширными административными, фискальными и судебными привилегиями.Остров Порту-Санту с гораздо меньшим количеством воды и растительности был передан третьему лорду-владельцу. Эта система лордов-собственников была введена в Бразилии в 1530-х годах.
В то время как сахар был королем, рабская часть населения архипелага составляла примерно 10 процентов. К 1460-м годам было подсчитано, что на Мадейре проживало две тысячи жителей. К началу шестнадцатого века здесь проживало двадцать тысяч человек, в том числе около двух тысяч рабов. Тем временем, чтобы поощрить колонизацию, главный остров Мадейра был разделен на два наследственных владения лордов, Фуншал и Машику, с обширными административными, фискальными и судебными привилегиями.Остров Порту-Санту с гораздо меньшим количеством воды и растительности был передан третьему лорду-владельцу. Эта система лордов-собственников была введена в Бразилии в 1530-х годах.
К началу шестнадцатого века Фуншал, столица Мадейры, был достаточно большим, с пятью тысячами жителей, чтобы получить статус города (1508 г.). Шесть лет спустя он стал резиденцией епархии (1514 г.) с юрисдикцией над всеми португальскими заморскими территориями. В это время архипелаг Мадейра продолжал оставаться важной промежуточной станцией для судов, идущих на Канарские острова и обратно, а также вдоль западного побережья Африки. К 1676 году население Мадейры достигло пятидесяти тысяч человек, из которых десять тысяч проживали в Фуншале.
К 1676 году население Мадейры достигло пятидесяти тысяч человек, из которых десять тысяч проживали в Фуншале.
Азорские острова, кажется, были открыты в 1427 году и были необитаемы. Два самых восточных острова (Санта-Мария и Сан-Мигель) Азорского архипелага находятся примерно в 840 милях от Португалии и в 420 милях от Мадейры. Сначала животных (особенно овец и коз) оставляли на необитаемых островах, чтобы заблудшие или потерпевшие кораблекрушение моряки имели пищу. К 1439 году было известно семь островов, в том числе средняя группа Терсейра, Фаял, Сан-Хорхе, Пику и Грасиоза, причем Терсейра находилась в семидесяти пяти морских милях от Сан-Мигеля.Наконец, два самых западных острова (Флорес и Корво), расположенные примерно в 1000 милях от Ньюфаундленда и в 375 милях к западу от Санта-Мария, были открыты около 1450 г. Диого де Тейве. Подход лорда-собственника также использовался на Азорских островах и, возможно, был даже более важным, чем на Мадейре. Древесина и вайда были первыми статьями экспорта, затем важное значение приобрела пшеница, хотя вайда и другие красители были основными статьями экспорта до конца семнадцатого века. Первоначальное заселение было медленным процессом, но к концу пятнадцатого века все девять островов Азорских островов были заселены поселенцами из Португалии, Фландрии и Мадейры.К 1500 году было пять городов. К 1550 году было два города — Ангра на Терсейре и Понта-Делгада в Сан-Мигеле — и двенадцать поселков. В 1534 году Ангра стала резиденцией епархии с юрисдикцией над всеми Азорскими островами.
Первоначальное заселение было медленным процессом, но к концу пятнадцатого века все девять островов Азорских островов были заселены поселенцами из Португалии, Фландрии и Мадейры.К 1500 году было пять городов. К 1550 году было два города — Ангра на Терсейре и Понта-Делгада в Сан-Мигеле — и двенадцать поселков. В 1534 году Ангра стала резиденцией епархии с юрисдикцией над всеми Азорскими островами.
В 1582 (на Сан-Мигеле) и 1583 (на Терсейре) войска дона Антонио (1531–1595), приора Крато и претендента на португальский престол, при поддержке французов потерпели поражение от испанца Альваро де Базана (1526). –1588), маркиз Санта-Крус. В период правления испанских Габсбургов (особенно в первые годы) Азорские острова часто подвергались нападениям со стороны английских, французских и голландских пиратов и корсаров.К 1587 г. население архипелага составляло тридцать три тысячи человек, а к 1695 г. число жителей оценивалось в восемьдесят пять тысяч. На протяжении шестнадцатого, семнадцатого и восемнадцатого веков большое количество избыточного населения Азорских островов и Мадейры мигрировало в португальские заморские колонии, особенно в Бразилию.
См. также Португалия .
БИБЛИОГРАФИЯ
Дункан, Т. Бентли. Атлантические острова: Мадейра, Азорские острова и Кабо-Вердес в торговле и мореплавании семнадцатого века. Чикаго и Лондон, 1972 г.
Оливейра Маркес, Антонио Энрике де. История Португалии. 2-е изд. New York, 1976.
Francis A. Dutra
Паттерны генетического разнообразия и структуры у видов, подвергшихся быстрой хромосомной радиации: аллозимный анализ домашних мышей с архипелага Мадейра
были отловлены в комменсальных местах обитания (дома, сады, фермы и возделываемые поля) с помощью живоловушек Шермана с приманкой из сардиновой пасты.Были проанализированы 158 мышей из 37 населенных пунктов Мадейры (экспедиции 1998 и 1999 гг.) вместе с 24 мышами из семи населенных пунктов Порту-Санту. Образцы с Мадейры произошли от различных хромосомных рас (см. Таблицу 1) и хромосомной гибридной зоны между расой Сантана и стандартными мышами в Фуншале, главном порту въезда на остров (см. Britton-Davidian et al., 2000, 2005). ). Кроме того, девять мышей были отловлены в двух местах в центральной части Португалии (Кайшас, 38°40 с.ш., 9°20 з.д. и Сантарен, 39°23 с.ш., 8°68 з.д.).Большинство этих местонахождений были объединены в соответствии с географической и/или хромосомной близостью в 22 выборки населения (17 на Мадейре, 4 в Порту-Санту и 1 в континентальной Португалии; см. рис. 1). Для сравнения образцы M. m. domesticus были пойманы вдоль восточного побережья южной Дании в мае 2000 г. (подробности см. в Smadja et al., 2004). Были отобраны несколько фермерских домов вокруг двух основных городов, в результате чего было получено 17 мышей в Зоммерстеде (Каства А, Рингтвед, Силлерупгаард, Каствра) и 21 мышь в Зондер Бьерте (Скартвед, Биндеруп, Зёндер Стендеруп, Лаурицминде).Названия островных рас Rb соответствуют номенклатуре, предложенной Piálek et al. (2005), за исключением того, что префикс P (для Португалии) был опущен для краткости. Таблица 1 Параметры генетического разнообразия для 33 изученных локусов в отобранных популяциях Рисунок 1
Britton-Davidian et al., 2000, 2005). ). Кроме того, девять мышей были отловлены в двух местах в центральной части Португалии (Кайшас, 38°40 с.ш., 9°20 з.д. и Сантарен, 39°23 с.ш., 8°68 з.д.).Большинство этих местонахождений были объединены в соответствии с географической и/или хромосомной близостью в 22 выборки населения (17 на Мадейре, 4 в Порту-Санту и 1 в континентальной Португалии; см. рис. 1). Для сравнения образцы M. m. domesticus были пойманы вдоль восточного побережья южной Дании в мае 2000 г. (подробности см. в Smadja et al., 2004). Были отобраны несколько фермерских домов вокруг двух основных городов, в результате чего было получено 17 мышей в Зоммерстеде (Каства А, Рингтвед, Силлерупгаард, Каствра) и 21 мышь в Зондер Бьерте (Скартвед, Биндеруп, Зёндер Стендеруп, Лаурицминде).Названия островных рас Rb соответствуют номенклатуре, предложенной Piálek et al. (2005), за исключением того, что префикс P (для Португалии) был опущен для краткости. Таблица 1 Параметры генетического разнообразия для 33 изученных локусов в отобранных популяциях Рисунок 1 Карты Мадейры и Порту-Санту. Цифры относятся к выбранным местам, а пунктирная линия указывает на места, объединенные по популяции и по расе. San=Santana, Asj=Arco di San Jorge, Pod=Ponta Delgada, Svi=Sao Vicente, Edc=Estreito de Calheta, Adc=Achadas da Cruz, Adc/Edc=гибридная популяция между расами Adc и Edc, Ldb=Lugar di Байшу, Пса = Порту-Санту, Фан = гибридная зона Фуншала.На вставке показано распределение населенных пунктов в Фуншале. Распределение слияний Rb между расами: San (включая Asj) = 2,19, 3,8, 4,16, 5,14, 6,7, 9,10, 11,12, 13,17, 15,18; Pod=3,8, 4,5, 9,18, 10,16, 11,12, 14,17; Svi=2,4, 3,8, 5,18, 6,7, 9,12, 10,16, 11,19, 13,17; Эдс=2,4, 3,14, 5,18, 6,7, 8,11, 9,12, 10,16, 13,17; Adc=2,4, 3,14, 5,18, 7,15, 8,11, 9,12, 10,16, 13,17; Лдб=2,4, 3,14, 6,7, 8,11, 9,12, 10,16, 13,17, 15,18.
Цифры относятся к выбранным местам, а пунктирная линия указывает на места, объединенные по популяции и по расе. San=Santana, Asj=Arco di San Jorge, Pod=Ponta Delgada, Svi=Sao Vicente, Edc=Estreito de Calheta, Adc=Achadas da Cruz, Adc/Edc=гибридная популяция между расами Adc и Edc, Ldb=Lugar di Байшу, Пса = Порту-Санту, Фан = гибридная зона Фуншала.На вставке показано распределение населенных пунктов в Фуншале. Распределение слияний Rb между расами: San (включая Asj) = 2,19, 3,8, 4,16, 5,14, 6,7, 9,10, 11,12, 13,17, 15,18; Pod=3,8, 4,5, 9,18, 10,16, 11,12, 14,17; Svi=2,4, 3,8, 5,18, 6,7, 9,12, 10,16, 11,19, 13,17; Эдс=2,4, 3,14, 5,18, 6,7, 8,11, 9,12, 10,16, 13,17; Adc=2,4, 3,14, 5,18, 7,15, 8,11, 9,12, 10,16, 13,17; Лдб=2,4, 3,14, 6,7, 8,11, 9,12, 10,16, 13,17, 15,18.
Кариотипы мышей с Мадейры, Порту-Санту, материковой Португалии и Дании были описаны ранее (Gazave et al., 2003; Матиас и др., 2004 г.; Смаджа и др., 2004; Бриттон-Дэвидиан и др., 2000, 2005; Нунес и др., 2005).
Аллозимный анализ
Аллозимный анализ был проведен на 229 мышах. В общей сложности 33 локуса были оценены в образцах печени, почек, сердца, плазмы и гемолизата с помощью электрофореза в крахмальном геле (см. Britton-Davidian, 1990 для ссылок на методы): алкогольдегидрогеназа ADH (1.1.1.1, Adh-1 ), альбумин ALB ( Alb-1 ), амилаза AMY (3.2.1.1, Amy-1 ), карбоангидраза CAR (4.2.1.1, Car-2 ), креатинкиназа CK (2.7.3.2, Ck-1 ), эстеразы ES (3.1.1.1, Es-1 , Es-2 , Es-3 , Es-10 , Es-15 ), гуаниндезаминаза GDA (3.5.4.3, Gda-1 ), α -глицерофосфатдегидрогеназа α GPD (1.1.059 GPD (1.1.059) 1.1.059 5 1.1.059 GPD (1.1.059) 1.1.059 GPD (1.1.059) , глиоксалаза GLO (4.4.1.5, Glo-1 ), амино-аспартаттрансаминазы GOT (2.6.1.1, Got-1 , Got-2 ), глицерофосфатизомераза GPI (5.3.1.9, Gpi-1 ), гидроксикислотная оксидаза HAO (1.1.3.15, Hao-1 ), гемоглобин HBB ( Hbb ), изоцитратдегидрогеназа IDH (1.1.1.42, Idh-1 , Idh 90 -2 ), лактатдегидрогеназа ЛДГ (1.
В общей сложности 33 локуса были оценены в образцах печени, почек, сердца, плазмы и гемолизата с помощью электрофореза в крахмальном геле (см. Britton-Davidian, 1990 для ссылок на методы): алкогольдегидрогеназа ADH (1.1.1.1, Adh-1 ), альбумин ALB ( Alb-1 ), амилаза AMY (3.2.1.1, Amy-1 ), карбоангидраза CAR (4.2.1.1, Car-2 ), креатинкиназа CK (2.7.3.2, Ck-1 ), эстеразы ES (3.1.1.1, Es-1 , Es-2 , Es-3 , Es-10 , Es-15 ), гуаниндезаминаза GDA (3.5.4.3, Gda-1 ), α -глицерофосфатдегидрогеназа α GPD (1.1.059 GPD (1.1.059) 1.1.059 5 1.1.059 GPD (1.1.059) 1.1.059 GPD (1.1.059) , глиоксалаза GLO (4.4.1.5, Glo-1 ), амино-аспартаттрансаминазы GOT (2.6.1.1, Got-1 , Got-2 ), глицерофосфатизомераза GPI (5.3.1.9, Gpi-1 ), гидроксикислотная оксидаза HAO (1.1.3.15, Hao-1 ), гемоглобин HBB ( Hbb ), изоцитратдегидрогеназа IDH (1.1.1.42, Idh-1 , Idh 90 -2 ), лактатдегидрогеназа ЛДГ (1. 1.1.27, Ldh-1 , Ldh-2 ), НАДФ-зависимые малатдегидрогеназы MOD (1.1.1.40, Mod-1 , Mod-2 ), NAD-зависимая малатдегидрогеназа MOR (1.1.1.37, Mor-1 , Mor-2 ), маннозофосфатизомераза MPI (5.3.1.8, Mpi-1 ), нуклеозидфосфорилаза NP (2.4.2.1, Np-1 ), фосфоглюкомутаза PGM (5.4.2.2, Pgm-1 , Pgm-2 ), пируваткиназа PK ( 2.7.1.40, Pk-1 , Pk-3 ) и супероксиддисмутазы СОД (1.15.1.1, Sod-1 ). Идентичность и номенклатуру аллелей оценивали путем сравнения с миграционными профилями аллелей, присутствующих у двух лабораторных линий мышей (C57Bl/6J и Balb/c; подробности процедуры см. в Britton-Davidian et al. (1989)).
1.1.27, Ldh-1 , Ldh-2 ), НАДФ-зависимые малатдегидрогеназы MOD (1.1.1.40, Mod-1 , Mod-2 ), NAD-зависимая малатдегидрогеназа MOR (1.1.1.37, Mor-1 , Mor-2 ), маннозофосфатизомераза MPI (5.3.1.8, Mpi-1 ), нуклеозидфосфорилаза NP (2.4.2.1, Np-1 ), фосфоглюкомутаза PGM (5.4.2.2, Pgm-1 , Pgm-2 ), пируваткиназа PK ( 2.7.1.40, Pk-1 , Pk-3 ) и супероксиддисмутазы СОД (1.15.1.1, Sod-1 ). Идентичность и номенклатуру аллелей оценивали путем сравнения с миграционными профилями аллелей, присутствующих у двух лабораторных линий мышей (C57Bl/6J и Balb/c; подробности процедуры см. в Britton-Davidian et al. (1989)).
Статистический анализ
Были оценены частоты аллелей, а также следующие показатели внутрипопуляционной изменчивости: среднее число аллелей ( A ), процент полиморфных локусов на образец (P%) и средняя ожидаемая гетерозиготность ( H e ), рассчитанных для небольших выборок с помощью программы BIOSYS-1 (Swofford, Selander, 1981). Все остальные параметры популяции были рассчитаны и протестированы в соответствии с GENEPOP v3.2a (Raymond and Rousset, 1995), если не указано иное.Во всех случаях использовались параметры по умолчанию. Точные тесты со связанными с ними значениями P были выполнены в каждой популяции для проверки отклонений от ожиданий Харди-Вайнберга и неравновесия по парному сцеплению между локусами. Генную дифференциацию между парами популяций оценивали с помощью точного критерия Фишера. Кроме того, были рассчитаны парные значения F ST с использованием оценщика θ , и их уровень значимости оценен с помощью тестов перестановки (1000) с использованием GENETIX v4.03 (Белхир и др., 1996–2004). Отношения между популяциями были исследованы путем построения дерева максимального правдоподобия (ML) из данных о частоте генов и дерева расстояний с использованием метода соседнего соединения (NJ). В качестве генетического расстояния использовался индекс Рейнольдса F ST , доступный в GENETIX v4.
Все остальные параметры популяции были рассчитаны и протестированы в соответствии с GENEPOP v3.2a (Raymond and Rousset, 1995), если не указано иное.Во всех случаях использовались параметры по умолчанию. Точные тесты со связанными с ними значениями P были выполнены в каждой популяции для проверки отклонений от ожиданий Харди-Вайнберга и неравновесия по парному сцеплению между локусами. Генную дифференциацию между парами популяций оценивали с помощью точного критерия Фишера. Кроме того, были рассчитаны парные значения F ST с использованием оценщика θ , и их уровень значимости оценен с помощью тестов перестановки (1000) с использованием GENETIX v4.03 (Белхир и др., 1996–2004). Отношения между популяциями были исследованы путем построения дерева максимального правдоподобия (ML) из данных о частоте генов и дерева расстояний с использованием метода соседнего соединения (NJ). В качестве генетического расстояния использовался индекс Рейнольдса F ST , доступный в GENETIX v4. 03. Оба дерева были построены с помощью пакета PHYLIP v3.5c (Felsenstein, 1985). Поддержка ветвей была установлена путем 500 повторных выборок начальной загрузки среди локусов. Графическое отображение древовидных топологий было нарисовано с помощью TreeView v1.6.6, распространяемое Пейджем (1996). Те же самые процедуры были применены к хромосомным данным, рассматривая каждое слияние Rb как локус с акроцентрическим и метацентрическим состояниями, рассматриваемыми как альтернативные аллели. В этом случае были проведены два дополнительных теста: неравновесное сцепление между слияниями Rb и аллозимными локусами и проверка корреляции между генетическими расстояниями (Nei, 1978) для слияний Rb и аллозимов с использованием процедуры Мантеля, реализованной в GENETIX v4.03. (1000 перестановок). P -значения для множественных тестов подвергали последовательной коррекции Бонферрони.
03. Оба дерева были построены с помощью пакета PHYLIP v3.5c (Felsenstein, 1985). Поддержка ветвей была установлена путем 500 повторных выборок начальной загрузки среди локусов. Графическое отображение древовидных топологий было нарисовано с помощью TreeView v1.6.6, распространяемое Пейджем (1996). Те же самые процедуры были применены к хромосомным данным, рассматривая каждое слияние Rb как локус с акроцентрическим и метацентрическим состояниями, рассматриваемыми как альтернативные аллели. В этом случае были проведены два дополнительных теста: неравновесное сцепление между слияниями Rb и аллозимными локусами и проверка корреляции между генетическими расстояниями (Nei, 1978) для слияний Rb и аллозимов с использованием процедуры Мантеля, реализованной в GENETIX v4.03. (1000 перестановок). P -значения для множественных тестов подвергали последовательной коррекции Бонферрони.
Для изучения генетического родства островных популяций с Португалией и Данией был проведен байесовский анализ структуры населения, реализованный в BAPSv. 4.14 (опция: анализ обученной смеси с использованием этих двух европейских образцов в качестве эталонного файла; Corander et al. , 2006). Чтобы сравнить генетическое разнообразие и родство популяций архипелага Мадейра с более крупной европейской выборкой, были использованы данные аллозимов еще 27 европейских и североафриканских популяций домашних мышей (Britton-Davidian et al., 1989; Britton-Davidian, 1990) были включены в анализ. H e , генетические дистанции и фенетические деревья были оценены по 31 локусу, общему для всех образцов ( Haox и Gda опущены). Различия в H e между географическими группами населения были проверены с помощью непараметрического критерия Краскела-Уоллиса. Генетическую структуру популяции исследовали с помощью анализов молекулярной дисперсии (AMOVA, SAMOVA), которые оценивают долю генной изменчивости на разных иерархических уровнях, используя информацию о географическом распределении частот аллозимов и попарном расстоянии между ними.
4.14 (опция: анализ обученной смеси с использованием этих двух европейских образцов в качестве эталонного файла; Corander et al. , 2006). Чтобы сравнить генетическое разнообразие и родство популяций архипелага Мадейра с более крупной европейской выборкой, были использованы данные аллозимов еще 27 европейских и североафриканских популяций домашних мышей (Britton-Davidian et al., 1989; Britton-Davidian, 1990) были включены в анализ. H e , генетические дистанции и фенетические деревья были оценены по 31 локусу, общему для всех образцов ( Haox и Gda опущены). Различия в H e между географическими группами населения были проверены с помощью непараметрического критерия Краскела-Уоллиса. Генетическую структуру популяции исследовали с помощью анализов молекулярной дисперсии (AMOVA, SAMOVA), которые оценивают долю генной изменчивости на разных иерархических уровнях, используя информацию о географическом распределении частот аллозимов и попарном расстоянии между ними. Было выполнено два подхода. Первый включал AMOVA (Excoffier et al., 1992) установленного пользователем иерархического географического расположения популяций в соответствии с кластерами, присутствующими в дереве Нью-Джерси. Процедура заключалась в пошаговом увеличении размера географических групп для выявления групп с максимальным расхождением между группами. Вторая серия тестов была реализована с использованием подхода вариантного пространственного анализа молекулярной дисперсии (SAMOVA) (Dupanloup et al., 2002), выявляющего группы популяций, максимально дифференцированные друг от друга.Как и в процедуре AMOVA, доля общей генной изменчивости, обусловленная различиями между группами популяций, максимальна в пределах последовательно уменьшающегося числа групп.
Было выполнено два подхода. Первый включал AMOVA (Excoffier et al., 1992) установленного пользователем иерархического географического расположения популяций в соответствии с кластерами, присутствующими в дереве Нью-Джерси. Процедура заключалась в пошаговом увеличении размера географических групп для выявления групп с максимальным расхождением между группами. Вторая серия тестов была реализована с использованием подхода вариантного пространственного анализа молекулярной дисперсии (SAMOVA) (Dupanloup et al., 2002), выявляющего группы популяций, максимально дифференцированные друг от друга.Как и в процедуре AMOVA, доля общей генной изменчивости, обусловленная различиями между группами популяций, максимальна в пределах последовательно уменьшающегося числа групп.
Планктонные гребневики архипелага Мадейра (северо-восточная Атлантика)
Агассис, А. (1865 г.) Североамериканские акалефы. Иллюстрированный каталог Музея сравнительной зоологии Гарвардского колледжа, 2, 1–234.
Алвес, Ф. , Каннинг-Клод, Дж., Рибейро, К., Гестозо, И. и Кауфманн, М.(2019) Местные бентосные сообщества на мелководных скалистых рифах находят убежище в морском охраняемом районе на острове Мадейра. Журнал прибрежной охраны, 23, 373–383. https://doi.org/10.1007/s11852-018-0669-y
, Каннинг-Клод, Дж., Рибейро, К., Гестозо, И. и Кауфманн, М.(2019) Местные бентосные сообщества на мелководных скалистых рифах находят убежище в морском охраняемом районе на острове Мадейра. Журнал прибрежной охраны, 23, 373–383. https://doi.org/10.1007/s11852-018-0669-y
Bigelow, H.B. (1904) Медузы с Мальдивских островов. Бюллетень Музея сравнительной зоологии Гарвардского колледжа, 39, 245–269.
Biscoito, M., Ribeiro, C. & Freitas, M. (2018) Аннотированный контрольный список рыб архипелага Мадейра (СВ Атлантика): I — Chondrichthyes. Zootaxa, 4429 (3), 459–494.https://doi.org/10.11646/zootaxa.4429.3.2
Бурсма, М., Мальзан, А.М., Греве, В. и Джавидпур, Дж. (2007) Первое появление гребневика Mnemiopsis leidyi в Северном море. Helgoland Marine Research, 61, 153–155. https://doi.org/10.1007/s10152-006-0055-2
Какабелос, Э., Гестосо, И., Рамальоса, П., Риера, Л., Нето, А.И. и Каннинг-Клод, Дж. (2019) Приливные комплексы на валунах и сплошных скалистых берегах: многомасштабный подход на субтропическом острове. Морское биоразнообразие, 49 (6), 2709–2723.https://doi.org/10.1007/s12526-019-01000-7
Морское биоразнообразие, 49 (6), 2709–2723.https://doi.org/10.1007/s12526-019-01000-7
Chi, X., Dierking, J., Hoving, H., Lüskow, F., Denda, A., Christiansen, B., Sommer, У., Хансен, Т. и Джавидпур, Дж. (2020) Борьба с желейной паутиной: трофическая экология студенистого зоопланктона в океанических пищевых сетях восточной тропической Атлантики, оцененная с помощью анализа стабильных изотопов. Лимнология и океанография, 66 (2), 289–305. https://doi.org/10.1002/lno.11605
Кристиансен, Б., Бухгольц, К., Бухгольц, Ф., Чи, X., Кристиансен, С., Денда, А., Хаусс, Х., Ховинг, Х.Й.Т., Янсен, С., Кауфманн, М., Кроншнабель, А., Люсков, Ф., Мартин, Б., Мертен, В., Силва, П., Пинейру, Н., Спрингер Б., Занкл С. и Зеймет Т. (2016) SEAMOX: Влияние подводных гор и зон минимума кислорода на пелагическую фауну в восточной части тропической Атлантики — Рейс № MSM49 — 28 ноября — 21 декабря 2015 г. — Лас Пальмас-де-Гран-Канария (Испания) — Минделу (Республика Кабо-Верде). МАРИЯ С. МЕРИАН-Берихте, MSM49, 1–42. https://doi.org/10.2312/cr_msm49
https://doi.org/10.2312/cr_msm49
Чун, К. (1879 г.) Die im Golf von Neapel erscheinenden Rippenquallen.Mittelmeer Zoologischen Station von Neapel zugleich repert Mittelmeerkd, 1, 180–217.
Чун, К. (1898) Die Ctenophoren der Plankton-Expedition. Ergebnisse der Plankton- Expedition der Humboldt-Stiftung, 2, 1–32.
Дейдун, А. (2011) Коллекция недавних наблюдений гребневиков на Мальтийских островах. Журнал Black Sea/Mediterranean Environment, 17 (1), 4–13.
Диас Бриз, Л., Санчес, Ф., Мари, Н., Мианзан, Х. и Джензано, Г. (2017) Желатиновый зоопланктон (гребневики, сальпы и медузы): важный пищевой ресурс рыб в умеренном поясе юго-западной части Атлантического океана Океан.Исследования морской биологии, 13 (6), 630–644. https://doi.org/10.1080/17451000.2016.1274403
Динаске, Дж., Тительман, Дж., Мёллер, Л.Ф., Сетала, О., Гранхаг, Л., Андерсен, Т., Бамстедт, У., Харальдссон, М., Хосиа, А., Катайисто, Т., Краг, Т., Купаринен, Дж., Шретер, М.Л., Сёндергаард, М., Тиселиус, П. и Риман, Л. (2012) Каскадные эффекты гребневика мнемиопсиса leidyi в планктонной пищевой сети в эстуарной системе с ограниченным содержанием питательных веществ. Серия «Прогресс морской экологии», 460, 49–61. https://дои.org/10.3354/meps09770
и Риман, Л. (2012) Каскадные эффекты гребневика мнемиопсиса leidyi в планктонной пищевой сети в эстуарной системе с ограниченным содержанием питательных веществ. Серия «Прогресс морской экологии», 460, 49–61. https://дои.org/10.3354/meps09770
Дутто, М.С., Джензано, Г.Н., Скиарити, А., Леканда, Дж., Хоффмайер, М.С. и Пратолонго, П.Д. (2017) Медузы и гребневики из эстуария Баия-Бланка и соседнего внутреннего шельфа (юго-западная часть Атлантического океана, Аргентина). Отчеты о морском биоразнообразии, 10, 14.
https://doi.org/10.1186/s41200-017-0114-1
Энгель-Соренсен, К., Андерсен, П. и Холмструп, М. (2009) Сохранение инвазивных гребневика Mnemiopsis leidyi с помощью кислого раствора Люголя. Журнал исследований планктона, 31 (8), 917–920.https://doi.org/10.1093/plankt/fbp030
Фрейтас, Р., Ромейрас, М., Силва, Л., Кордейро, Р., Мадейра, П., Гонсалес, Х.А., Виртц, П., Фалькон, Дж.М., Брито, А., Флотер, С.Р., Афонсо, П., Протейро, Ф., Вьера-Родригес, М.А., Нето, А.И., Харун, Р. , Фарминьяо, Дж.Н., Ребело, А.С., Баптиста, Л., Мело , К.С., Мартинес, А., Нуньес, Дж., Бернинг, Б., Джонсон, М.Е. и Авила, С.П. (2019) Реструктуризация биогеографической единицы «Макаронезии»: морской мультитаксонный биогеографический подход. Научные отчеты, 9, 15792.https://doi.org/10.1038/s41598-019-51786-6
, Фарминьяо, Дж.Н., Ребело, А.С., Баптиста, Л., Мело , К.С., Мартинес, А., Нуньес, Дж., Бернинг, Б., Джонсон, М.Е. и Авила, С.П. (2019) Реструктуризация биогеографической единицы «Макаронезии»: морской мультитаксонный биогеографический подход. Научные отчеты, 9, 15792.https://doi.org/10.1038/s41598-019-51786-6
Gegenbaur, C. (1856 г.) Studien über Organization und Systematik der Ctenophoren. Archiv für Naturgeschichte, XXII, 163–205
Гиббонс, М.Дж., Пикша, С.Х.Д., Мацумото, Г.И. и Фостер, К. (2021) Находки гребневиков из Южной Африки. PeerJ, 9, e10697. https://doi.org/10.7717/peerj.10697
Гирибет Г. и Эджкомб Г. (2020) Древо жизни беспозвоночных. Издательство Принстонского университета, Принстон, Нью-Джерси, 589 стр. https://doi.org/10.1515/9780691197067
Greve, W. (1975) Ctenophora. Fiche d’Identification du Zooplankton, 146, 6–11.
Gueroun, SKM, Javidpour, J., Andrade, C., Nogueira, N., Freitas, M. & Canning-Clode, J. (2021) Модели и тенденции разнообразия пелагических Cnidaria и Ctenophora в островных системах Macaronesia (СВ Атлантика) . Морское биоразнообразие, 51, 32. https://doi.org/10.1007/s12526-021-01174-z
Морское биоразнообразие, 51, 32. https://doi.org/10.1007/s12526-021-01174-z
Пикша, С.Х.Д. (2007) Сравнительное пищевое поведение планктонных гребневиков. Интегративная и сравнительная биология, 47 (6), 847–853.https://doi.org/10.1093/icb/icm088
Ханссон, Х. Г. (2006) Гребневики Балтийского и прилегающих морей – захватчик Мнемиопсис уже здесь! Водные вторжения, 1 (4), 295–298. https://doi.org/10.3391/ai.2006.1.4.16
Harbison, G.R. (1985) К изучению биогеографии пелагических гребневиков. Материалы международной конференции, Нидерланды, 1985 г., стр. 112–117.
Харбисон Г.Р., Мадин Л.П. и Суонберг Н.Р. (1978) О естественной истории и распространении океанических гребневиков.Deep-Sea Research, 25 (3), 237–256. https://doi.org/10.1016/0146-6291(78)-8
Harbison, G.R. и Миллер Р.Л. (1986) Не все гребневики являются гермафродитами. Исследования по систематике, распространению, половой жизни и развитию двух видов Ocyropsis. Морская биология, 90, 413–424. https://doi. org/10.1007/BF00428565
org/10.1007/BF00428565
Хидака М., Нишикава Дж. и Линдси Д.Дж. (2021) Желатиновое сообщество зоопланктона вокруг гидротермально активной глубоководной кальдеры: результаты видеозаписей ROV.Исследования планктона и бентоса, 16 (1), 40–58. https://doi.org/10.3800/pbr.16.40
Кауфманн М.Дж., Сантос Ф. и Маранхао М. (2015 г.) Контрольный список нано- и микрофитопланктона у острова Мадейра (северо-восточная Атлантика) с некоторыми историческими примечаниями. Новая Хедвигия, 101, 205–232. https://doi.org/10.1127/nova_hedwigia/2015/0265
Knowler, D. (2005) Переоценка стоимости биологического вторжения: Mnemiopsis leidyi в Черном море. Экологическая экономика, 52, 187–199. https://doi.org/10.1016/j.ecolecon.2004.06.013
Лесюер, Калифорния (1813) Mémoire sur quelques nouvelles espèces d’animaux mollusques et radiaires recueillis dans la Méditerranée près de Nice. Nouveau Bulletin des Sciences par la Société Philomatique de Paris, Series 2, 3 (69), 281–285
Licandro, P., Blackett, M. , Fischer, A., Hosia, A., Kennedy, J., Kirby, Р.Р., Рааб К., Стерн Р. и Трантер П. (2015) Биогеография медуз в Северной Атлантике традиционными и геномными методами. Научные данные о системе Земли, 7, 173–191.https://doi.org/10.5194/essd-7-173-2015
, Fischer, A., Hosia, A., Kennedy, J., Kirby, Р.Р., Рааб К., Стерн Р. и Трантер П. (2015) Биогеография медуз в Северной Атлантике традиционными и геномными методами. Научные данные о системе Земли, 7, 173–191.https://doi.org/10.5194/essd-7-173-2015
Линдси, Д.Дж., Уметсу, М., Гроссманн, М., Мияке, Х. и Ямамото, Х. (2015) Желатиновое сообщество макропланктона в Гидротермальный источник Хатома-Нолл. В: Ишибаши Дж., Окино К. и Сунамура М. (ред.), Биосфера морского дна, связанная с гидротермальными системами: концепция TAIGA. Спрингер, Токио, стр. 639–666. https://doi.org/10.1007/978-4-431-54865-2_51
Лонгхерст, А., Сатиендранат, С., Платт, Т. и Каверхилл, К. (1995) Оценка глобального первичного производства в океана по данным спутникового радиометра.Журнал исследований планктона, 17, 1245–1271. https://doi.org/10.1093/plankt/17.6.1245
Марторелли, С.Р. (2001) Digenea паразиты медуз и гребневиков южной Атлантики. Гидробиология, 451, 305–310. https://doi.org/10.1023/A:1011862406670
Mayer, AG (1912) Гребневики атлантического побережья Северной Америки. Институт Карнеги, Вашингтон, 58 стр. https://doi.org/10.5962/bhl.title.5968
Институт Карнеги, Вашингтон, 58 стр. https://doi.org/10.5962/bhl.title.5968
Mianzan, H.W. (1999) Гребневики. В: Деметрио, Б. (ред.), Зоопланктон Южной Атлантики.Том 1. Backhuys Publishers, Лейден, стр. 561–573.
Migotto, AE & Marques, AC (2007) Идентификационный ключ для гребневиков с побережья Бразилии. Biota Neotropica, 7 (3), 242–350.
Миллс, К.Э. и Хэддок, С.Х.Д. (2007) Ключ к гребневикам. В: Карлтон, Дж.Т. (Ред.), Руководство Лайта и Смита по литоральным беспозвоночным от Центральной Калифорнии до Орегона, полностью переработанное и дополненное. University of California Press, Окленд, Калифорния, стр. 189–199. https://doi.org/10.1525/9780520930438-015
Моро, Л., Боэлькхе, В., Эскаттлар, Дж. и Бакалладо, Дж.Дж. (2013) Nota sobre la presencia de Ocyropsis crystallina (Rang, 1828) (Ctenophora: Lobata) на Канарских островах. Revista de la Academia Canaria de Ciencias, 25, 129–133.
Мюллер, О.Ф. (1776) Zoologiæ Danicæ Prodromus, seu Animalium Daniae et Norvegiæ indigenarum characteres, nomina, et synonyma imprimis Popularium. typis Hallageriis, Havniæ [Копенгаген], xxxii + 274 стр. https://doi.org/10.5962/bhl.title.63795
typis Hallageriis, Havniæ [Копенгаген], xxxii + 274 стр. https://doi.org/10.5962/bhl.title.63795
Narciso, Á., Caldeira, R., Reis, J., Хоппенрат, М., Кашао, М. и Кауфманн, М. (2019) Влияние временной фронтальной зоны на пространственное распределение существующих кокколитофоридов вокруг архипелага Мадейра (северо-восточная Атлантика). Эстуарий, прибрежная и шельфовая наука, 223, 25–38. https://doi.org/10.1016/j.ecss.2019.04.014
Оцука С., Койке К., Линдси Д., Нисикава Дж., Мияке Х., Кавахара М., Муляди, Муджионо Н., Хироми Дж. и Комацу Х. (2009) Симбионты морских медуз и гребневиков. Исследования планктона и бентоса, 4, 1–13.https://doi.org/10.3800/pbr.4.1
Oliveira, O.M.P. и Migotto, AE (2006) Пелагические гребневики из канала Сан-Себастьян, юго-восток Бразилии. Zootaxa, 1183 (1), 1–26. https://doi.org/10.11646/zootaxa.1183.1.1
Оливейра, ОМП, Миранда, Т.П., Араужо, Э.М., Айон, П., Седеньо-Поссо, К.М., Сепеда-Меркадо, А.А., Кордова, П., Кунья, А.Ф., Джензано, Г.Н., Хаддад, М. А., Мианзан, Х.В., Миготто, А.Е., Миранда, Л.С., Морандини, А.С., Нагата, Р.М., Насименто, К.Б., Ногейра, М., Пальма, С., Киньонес Дж., Родригес К.С., Скарабино Ф., Скиарити А., Стампар С.Н., Тронолон В.Б. и Маркес, AC (2016) Перепись Cnidaria (Medusozoa) и Ctenophora из морских вод Южной Америки. Zootaxa, 4194 (1), 1–256. https://doi.org/10.11646/zootaxa.4194.1.1
А., Мианзан, Х.В., Миготто, А.Е., Миранда, Л.С., Морандини, А.С., Нагата, Р.М., Насименто, К.Б., Ногейра, М., Пальма, С., Киньонес Дж., Родригес К.С., Скарабино Ф., Скиарити А., Стампар С.Н., Тронолон В.Б. и Маркес, AC (2016) Перепись Cnidaria (Medusozoa) и Ctenophora из морских вод Южной Америки. Zootaxa, 4194 (1), 1–256. https://doi.org/10.11646/zootaxa.4194.1.1
Purcell, J.E. (2005) Влияние климата на образование медуз и цветение гребневиков: обзор. Журнал Морской биологической ассоциации Великобритании, 85, 461–476. https://doi.org/10.1017/S0025315405011409
Рамальоса, П., Гестосо, И., Роча, Р.М., Ламберт, Г. и Каннинг-клоуд, Дж. (2021) Биоразнообразие асцидий на мелководье архипелага Мадейра: исследования обрастания искусственных субстратов и новые записи. Региональные исследования в области морских наук, 43, 101672. https://doi.org/10.1016/j.rsma.2021.101672
Rang, S. (1827) Описание нового жанра класса Acalèphes. Bulletin d’histoire naturelle de la Socieìteì Linneìenne de Bordeaux, 1 (4), 316–319
Rankin, J. J. (1956) Структура и биология Vallicula multiformis, gen.и др. nov., платиктенидный гребневик. Журнал Лондонского Линнеевского общества, Зоология, 43 (289), 55–71. https://doi.org/10.1111/j.1096-3642.1956.tb02507.x
J. (1956) Структура и биология Vallicula multiformis, gen.и др. nov., платиктенидный гребневик. Журнал Лондонского Линнеевского общества, Зоология, 43 (289), 55–71. https://doi.org/10.1111/j.1096-3642.1956.tb02507.x
Рибейро, К., Араужо, А.Дж., Бискойто, М. и Фрейтас, М. (2005) Рыбные сообщества залива Кайш-ду-Карван (Мадейра) о-в) определяется методом визуального учета. Журнал биологии рыб, 67, 1568–1584. https://doi.org/10.1111/j.1095-8649.2005.00861.x
Сала, И., Калдейра, Р.М., Эстрада-Аллис, С.Н., Фруфе, Э. и Кувелар, X.(2013) Лагранжевы транспортные пути в северо-восточной Атлантике и их влияние на окружающую среду. Лимнология и океанография: жидкости и окружающая среда, 3, 40–60. https://doi.org/10.1215/21573689-2152611
Сан-Мартин, Г., Альварес-Кампос, П., Кондо, Ю., Нуньес, Дж., Фернандес-Аламо, М.А., Плейхель, Ф., Гетц, FE, Nygren, A. & Osborn, K. (2021) Новая симбиотическая ассоциация морских кольчатых червей: эктопаразиты гребенчатых желе. Зоологический журнал Линнеевского общества, 191, 672–694. https://doi.org/10.1093/zoolinnean/zlaa034
https://doi.org/10.1093/zoolinnean/zlaa034
Сарс, М. (1856) Fauna littoralis Norvegiae, oder Beschreibung und Abbildungen neuer oder oder wenig bekannten Seethiere. Фредерик Д. Бейер, Берген, 144 стр.
Салливан, Б.К., Керен, Д. Ван и Клэнси, М. (2001) Сроки и размер цветения гребневика Mnemiopsis leidyi в зависимости от температуры в заливе Наррагансетт, Род-Айленд. В: Purcell J.E., Graham, W.M. и Дюмон, Х. Дж. (редакторы), Цветение медуз: экологическое и социальное значение. Спрингер, Дордрехт, стр. 113–120.https://doi.org/10.1023/A:1011848327684
Sweetman, A.K. & Chapman, A. (2015) Первая оценка скорости потока туш медуз (падений медуз) в бентос показывает важность студенистого материала для биологического C-цикла в экосистемах с преобладанием медуз. Frontiers in Marine Science, 2, 47. https://doi.org/10.3389/fmars.2015.00047
Sweetman, A.K., Smith, C.R., Dale, T. & Jones, D.O.B. (2014) Быстрое поедание туш медуз показывает важность студенистого материала для глубоководных пищевых сетей. Proceedings of the Royal Society B, 281 (1796), 2014–2210. https://doi.org/10.1098/rspb.2014.2210
Proceedings of the Royal Society B, 281 (1796), 2014–2210. https://doi.org/10.1098/rspb.2014.2210
Thibault-botha, D. & Bowen, T. (2004) Влияние консервации формалина на Pleurobrachia bachei ( Гребневики). Журнал экспериментальной морской биологии и экологии, 303, 11–17. https://doi.org/10.1016/j.jembe.2003.10.017
Wirtz, P. (1998) Двенадцать беспозвоночных и восемь видов рыб, новых для морской фауны Мадейры, и обсуждение зоогеографии этого района. Helgolander Meeresuntersuchungen, 52, 197–207.https://doi.org/10.1007/BF028
Wirtz, P. (2007) О коллекции гидроидов (Cnidaria, Hydrozoa) с архипелага Мадейра. Архипелаг — Науки о жизни и море, 24, 11–16.
Виртц, П., Фирке, Р. и Бискойто, Дж. (2008) Прибрежные рыбы острова Мадейра – новые записи и аннотированный контрольный список. Zootaxa, 1715 (1), 1–26. https://doi.org/10.11646/zootaxa.1715.1.1
границ | Архипелаг Мадейра как значительный источник разнообразия актиномицетов морского происхождения с противораковым и противомикробным потенциалом
Введение
Морские актиномицеты оказались важным источником новых вторичных метаболитов, а также многообещающим источником фармацевтически важных агентов (Magarvey et al. , 2004; Дженсен и др., 2005a,b; Лам, 2006 г.; Прюдом и др., 2008 г.; Асолкар и др., 2009; Рахман и др., 2011; Субрамани и Алберсберг, 2012). Было показано, что производство разнообразных вторичных метаболитов актиномицетами морского происхождения даже выше, чем у наиболее плодовитого природного источника биоактивных вторичных метаболитов на сегодняшний день: рода Streptomyces (Dharmaraj, 2010). Например, представители морского облигатного рода Salinispora , которые оказались особенно богатым источником новых химических структур и новых классов соединений (Jensen and Mafnas, 2006), в том числе некоторые из них прошли фазу II клинических испытаний рака (Cragg и другие., 2014).
, 2004; Дженсен и др., 2005a,b; Лам, 2006 г.; Прюдом и др., 2008 г.; Асолкар и др., 2009; Рахман и др., 2011; Субрамани и Алберсберг, 2012). Было показано, что производство разнообразных вторичных метаболитов актиномицетами морского происхождения даже выше, чем у наиболее плодовитого природного источника биоактивных вторичных метаболитов на сегодняшний день: рода Streptomyces (Dharmaraj, 2010). Например, представители морского облигатного рода Salinispora , которые оказались особенно богатым источником новых химических структур и новых классов соединений (Jensen and Mafnas, 2006), в том числе некоторые из них прошли фазу II клинических испытаний рака (Cragg и другие., 2014).
Различное давление окружающей среды может заставить бактерии адаптироваться к определенному месту. Эти бактерии могут содержать специфические гены, ответственные за синтез больших мультиферментных соединений, таких как поликетиды и нерибосомальные пептиды, или их гибриды (Penn et al., 2009; Bose et al.
 Таким образом, культивирование новых видов бактерий из разных мест океана остается продуктивным подходом к открытию природных продуктов (Becerril-Espinosa et al., 2013). Было исследовано разнообразие актиномицетов в отложениях, собранных в Тихом океане и Карибском море (Jensen, Lauro, 2008; Solano et al., 2009; Prieto-Davó et al., 2013). Более 500 сортов Actinobacteria были выделены из морских отложений Палау (Gontang et al., 2007), а роды актиномицетов, выделенные из отложений Марианской впадины, включают Dermacoccus, Kocuria, Micromonospora, Streptomyces, Tsukammurella и Williamsia (Pathom-aree et al., 2006). Однако разнообразие актиномицетов из отложений Атлантического океана практически не изучено (Weyland, 1969; Walker and Colwell, 1975; Williams et al., 1999; Fiedler et al., 2005), и некоторые исследования по изоляции привели к выделению нескольких родов, включая Nocardia, Micromonospora, Microbispora и Streptomyces , из отложений, собранных в Чесапикском заливе и Северном море (Weyland, 1969, 1984; Takizawa et al.
Таким образом, культивирование новых видов бактерий из разных мест океана остается продуктивным подходом к открытию природных продуктов (Becerril-Espinosa et al., 2013). Было исследовано разнообразие актиномицетов в отложениях, собранных в Тихом океане и Карибском море (Jensen, Lauro, 2008; Solano et al., 2009; Prieto-Davó et al., 2013). Более 500 сортов Actinobacteria были выделены из морских отложений Палау (Gontang et al., 2007), а роды актиномицетов, выделенные из отложений Марианской впадины, включают Dermacoccus, Kocuria, Micromonospora, Streptomyces, Tsukammurella и Williamsia (Pathom-aree et al., 2006). Однако разнообразие актиномицетов из отложений Атлантического океана практически не изучено (Weyland, 1969; Walker and Colwell, 1975; Williams et al., 1999; Fiedler et al., 2005), и некоторые исследования по изоляции привели к выделению нескольких родов, включая Nocardia, Micromonospora, Microbispora и Streptomyces , из отложений, собранных в Чесапикском заливе и Северном море (Weyland, 1969, 1984; Takizawa et al. , 1993). Более поздние исследования выявили очень высокую численность Micromonospora и Streptomyces в Тронхейм-фьорде, Норвегия (Bredholt et al., 2008), в то время как некоторые исследования отложений в Нью-Брансуике, Канада, зависящие от культивирования, выявили присутствие Micromonospora, Nocardia. , Nocardiopsis и Pseudonocardia , в дополнение к роду Streptomyces (Duncan et al., 2014, 2015). Другие места в океане, такие как глубокие отложения региона Макаронезии, на которые оказывает влияние селевой поток Сахары, были хорошим источником родов Streptomyces и Rhodococcus (Stach et al., 2003).
, 1993). Более поздние исследования выявили очень высокую численность Micromonospora и Streptomyces в Тронхейм-фьорде, Норвегия (Bredholt et al., 2008), в то время как некоторые исследования отложений в Нью-Брансуике, Канада, зависящие от культивирования, выявили присутствие Micromonospora, Nocardia. , Nocardiopsis и Pseudonocardia , в дополнение к роду Streptomyces (Duncan et al., 2014, 2015). Другие места в океане, такие как глубокие отложения региона Макаронезии, на которые оказывает влияние селевой поток Сахары, были хорошим источником родов Streptomyces и Rhodococcus (Stach et al., 2003).
В этом исследовании для культивирования разнообразных морских актиномицетов с архипелага Мадейра использовался подход, зависящий от культивирования. Основными целями этой работы было исследование бактериального разнообразия в неизведанном месте в Атлантическом океане и поиск новых актиномицетов морского происхождения с потенциальной биологической активностью. Результаты выявили значительное разнообразие актиномицетов в этом регионе Атлантического океана, включая штаммы, которые могут представлять новые виды или новые экотипы, принадлежащие к ранее описанным морским группам.Кроме того, наше исследование подтверждает фармацевтический потенциал актиномицетов морского происхождения из неисследованных областей океанского дна.
Результаты выявили значительное разнообразие актиномицетов в этом регионе Атлантического океана, включая штаммы, которые могут представлять новые виды или новые экотипы, принадлежащие к ранее описанным морским группам.Кроме того, наше исследование подтверждает фармацевтический потенциал актиномицетов морского происхождения из неисследованных областей океанского дна.
Материалы и методы
Коллекция образцов
Шестьсот шестьдесят две пробы океанических отложений были отобраны в атлантическом экорегионе Макаронезия, у берегов архипелага Мадейра, в южных пределах Мадейры (24 станции) и островов Порту-Санту (15 станций) и в западной части островов Дезертас ( 6 станций) с 4 по 17 июня 2012 года.Образцы состояли из 662 отложений, чередующихся от мелкозернистого ила до мелких камней и мелких кусочков мертвого коралла. Мелководные отложения собирались подводным плаванием с глубин 1–20 м. Глубоководные отложения были собраны с помощью модифицированного пробоотборника отложений (Kahlsico, El Cajon, CA, модель № 214WA110), который не требует прикрепления к лебедке и может достигать глубины до 1310 м. Каждый образец переносили в маркированный стерильный пакет (Nasco Whirl-pack) и немедленно хранили на льду для транспортировки на берег.Долгосрочное хранение было при температуре -20°С.
Каждый образец переносили в маркированный стерильный пакет (Nasco Whirl-pack) и немедленно хранили на льду для транспортировки на берег.Долгосрочное хранение было при температуре -20°С.
Методы выделения и культивирования
662 образца осадка были обработаны с использованием следующего метода сушки: осадок, ранее хранившийся при температуре -20°C в течение 1 месяца, сушили в течение ночи в ламинарном боксе и, когда происходило комкование, слегка растирали пестиком в стерилизованной спиртом ступке. Автоклавированную пробку (диаметром 1–2 см) прижимали к осадку, а затем многократно прижимали к поверхности агаровой пластины по часовой стрелке, создавая эффект серийного разведения.Этот метод предназначен для уменьшения числа грамотрицательных бактерий и обогащения медленнорастущими спорообразующими актиномицетами. Образцы обрабатывали и инокулировали, как описано выше, на поверхность одной-трех следующих агаровых сред. Все среды готовили из 1 л природной морской воды и деионизированной воды в соотношении 75:25 и содержали противогрибковый агент циклогексимид и w (100 мг/мл -1 ). Среда 1 (А1) 18 г агара, 10 г крахмала, 4 г дрожжевого экстракта, 2 г пептона, среда 2 (1/2 А1) 18 г агара, 5 г крахмала, 2 г дрожжевого экстракта, 1 г пептона и среды 3 (SWA) 18 г агара.
Среда 1 (А1) 18 г агара, 10 г крахмала, 4 г дрожжевого экстракта, 2 г пептона, среда 2 (1/2 А1) 18 г агара, 5 г крахмала, 2 г дрожжевого экстракта, 1 г пептона и среды 3 (SWA) 18 г агара.
Количественное определение и выделение актиномицетов
Инокулированные чашки Петри инкубировали при комнатной температуре ( c. 25–28°C) и периодически в течение 6 месяцев контролировали рост актиномицетов. Количество актиномицетов определяли на каждой чашке на глаз. Актиномицеты распознавали по наличию нитевидных гиф и/или по образованию жестких кожистых колоний, прилипших к поверхности агара. Таким образом, в исследование были включены только мицелиеобразующие бактерии порядка Actinomycetales .Только в 198 пробах были обнаружены мицелиеобразующие бактерии. Для каждой чашки, на которой образовались колонии актиномицетов, подсчитывали общее количество наблюдаемых колоний и получали представителей всех морфотипов в чистой культуре путем многократного пересева из одной колонии. 421 из этих колоний последовательно переносили на новые среды до получения чистых культур. Четыреста восемь штаммов актиномицетов выращивали в жидкой культуре (среда 1 без агара) и криоконсервировали при -80°С в 10% глицерине.Таксономическая классификация была выполнена с помощью анализа секвенирования 16S рРНК, и выделенные штаммы были проверены на потребность в морской воде для роста (см. ниже).
421 из этих колоний последовательно переносили на новые среды до получения чистых культур. Четыреста восемь штаммов актиномицетов выращивали в жидкой культуре (среда 1 без агара) и криоконсервировали при -80°С в 10% глицерине.Таксономическая классификация была выполнена с помощью анализа секвенирования 16S рРНК, и выделенные штаммы были проверены на потребность в морской воде для роста (см. ниже).
Влияние морской воды на рост
Все 408 выделенных актиномицетов были протестированы на потребность в морской воде для роста, т.е. обязательная морская оценка. Это достигается путем замены морской воды деионизированной водой в питательной среде и наблюдения за влиянием на рост (Jensen et al., 2005b). Используя стерильную петлю, клетки из четко определенной колонии удаляли с чашки А1, приготовленной с натуральной морской водой, и высевали штрихом на чашки А1, приготовленные с морской водой и деионизированной водой.Планшеты инкубировали при 25-28°С и наблюдали за ростом в течение 6 месяцев.
Экстракция ДНК, амплификация гена 16S рРНК, секвенирование и таксономическая идентификация
Все 421 чистый штамм были первоначально сгруппированы на основе наличия или отсутствия воздушного мицелия, размера колонии, морфологии, цвета колонии, внешнего вида спор, образования диффундирующего пигмента, а также наличия или отсутствия воздушных гиф и влияния морской воды на рост.Для секвенирования гена 16S рРНК и филогенетического анализа были отобраны представители каждой группы. Штаммы культивировали в 4 мл среды А1, встряхивая при 200 об/мин и 25°С в течение 3–7 сут. Набор Wizard ® для очистки геномной ДНК (Promega, Мэдисон, Висконсин, США) из протокола грамположительных бактерий использовали, как описано в Техническом руководстве Wizard ® для очистки геномной ДНК, № TM050, с небольшими изменениями. Мы обнаружили, что более длительные инкубационные периоды литического фермента (т. э., лизоцим) и раствор РНКазы были необходимы для получения эффективных количеств геномной ДНК. Ген 16S рРНК был амплифицирован в полимеразной цепной реакции (ПЦР) с использованием праймеров 27F (5′-AGAGTTTGATCCTGGCTCAG-3′) и 1492R (5′-TACGGCTACCTTGTTACGACTT-3′) (Gontang et al., 2007) и продуктов, очищенных с помощью SureClean. Набор для очистки ПЦР (BioLine, Лондон, Великобритания) с использованием протокола, предоставленного производителем. Очищенные реакции ПЦР секвенировали в цикле с перечисленными выше праймерами в STAB VIDA, Lda (http://www.stabvida.net/), с использованием набора для секвенирования цикла ABI BigDye ® Terminator v3.1 и очищенный. Очищенные продукты анализировали на генетическом анализаторе ABI PRISM ® 3730xl, а следы последовательности редактировали с помощью Sequencing Analysis 5.3.1 от Applied. Все полученные последовательности сравнивали с базой данных GenBank по алгоритму blastn.
э., лизоцим) и раствор РНКазы были необходимы для получения эффективных количеств геномной ДНК. Ген 16S рРНК был амплифицирован в полимеразной цепной реакции (ПЦР) с использованием праймеров 27F (5′-AGAGTTTGATCCTGGCTCAG-3′) и 1492R (5′-TACGGCTACCTTGTTACGACTT-3′) (Gontang et al., 2007) и продуктов, очищенных с помощью SureClean. Набор для очистки ПЦР (BioLine, Лондон, Великобритания) с использованием протокола, предоставленного производителем. Очищенные реакции ПЦР секвенировали в цикле с перечисленными выше праймерами в STAB VIDA, Lda (http://www.stabvida.net/), с использованием набора для секвенирования цикла ABI BigDye ® Terminator v3.1 и очищенный. Очищенные продукты анализировали на генетическом анализаторе ABI PRISM ® 3730xl, а следы последовательности редактировали с помощью Sequencing Analysis 5.3.1 от Applied. Все полученные последовательности сравнивали с базой данных GenBank по алгоритму blastn.
Регистрационные номера последовательностей нуклеотидов
Все нехимерные нуклеотидные последовательности из 400 культивируемых штаммов актиномицетов, о которых сообщалось в этом исследовании, были депонированы в GenBank под регистрационными номерами KT446002–KT446401, KJ685362–KJ685387 и KP869059–KP869064, доступными на http://www. ncbi.nlm.nih.gov/genbank (дополнительная таблица 1).
ncbi.nlm.nih.gov/genbank (дополнительная таблица 1).
Группы операционных таксономических единиц (OTU)
OTU были созданы с использованием 400 частичных последовательностей 16S рРНК (~350 п.н.) из культивируемых актиномицетов с идентичностью 97% с использованием команды cluster.classic в программе MOTHUR (http://www.mothur.org/), которая сравнивает последовательности алгоритмом «среднего соседа» (Schloss et al., 2009). Репрезентативная последовательность из каждой OTU была получена с использованием команды get.oturep в MOTHUR, и были получены почти полные последовательности (~ 1300 п.н.) для этих 71 представителя.Эти репрезентативные последовательности сравнивали с базой данных GenBank, чтобы определить их таксономическую принадлежность, а также процентную идентичность с 16S рРНК из их ближайшей зарегистрированной последовательности (соседней).
Анализы оценки разрежения и разнообразия
Четыреста частичных последовательностей были использованы для анализа разрежения и оценки разнообразия с использованием непараметрического статистического оценщика Chao1 с командой reefaction. single от MOTHUR.
single от MOTHUR.
Новое определение OTU
репрезентативных штамма сравнивали с типовыми штаммами в базе данных EZTaxon на предмет сходства последовательностей 16S рРНК. Когда последовательности были <98% сходны с ближайшим типовым штаммом, все штаммы в этой OTU затем сравнивали с типовым штаммом, чтобы определить, можно ли всю OTU считать новой или нет. Мы использовали сходство 98% для проведения более строгого анализа при принятии решения о новизне штаммов.
Филогенетический анализ
71 полная последовательность и их ближайшие соседние последовательности GenBank были выровнены с использованием ClustalX, и это выравнивание использовалось для создания филогенетического дерева максимального правдоподобия из 1279 п.н. с использованием MEGA версии 6.06 (http://www.megasoftware.net) с 1000 загрузчиками (Tamura et al., 2013).
Анализ UniFrac
Изолированные актиномицеты с архипелага Мадейра были разделены на мелкие (≤ 100 м) и глубокие (> 100 м) среды, и был проведен анализ UniFrac, чтобы выявить любые филогенетические различия между этими средами (Lozupone and Knight, 2005). При этом образцы были разделены по характеристикам осадка и построена дендограмма несходства UniFrac.UniFrac использует информацию о присутствии и количестве от OTU в сочетании с филогенетическим деревом для выполнения этих сравнений. Все анализы, выполненные на UniFrac, были взвешены и нормализованы для получения более надежного результата.
При этом образцы были разделены по характеристикам осадка и построена дендограмма несходства UniFrac.UniFrac использует информацию о присутствии и количестве от OTU в сочетании с филогенетическим деревом для выполнения этих сравнений. Все анализы, выполненные на UniFrac, были взвешены и нормализованы для получения более надежного результата.
Приготовление и фракционирование сырых экстрактов бактерий
Всего было выделено 400 штаммов актиномицетов, культивированных в идентичных условиях роста. Они заключались в выращивании каждого штамма в 90 мл среды М1 при встряхивании при 220 об/мин при 25°С в течение 7 дней. Посевные культуры (5 мл в колбах на 50 мл, среда 1) выращивали в течение 7 дней до переноса.Культуральный бульон экстрагировали EtOAc. Органический слой концентрировали досуха в вакууме и остаток повторно суспендировали в ДМСО при концентрации 10 мг/мл -1 .
Противораковый скрининг в клетках карциномы толстой кишки человека HCT116
Культура клеток
Клеточная линия карциномы толстой кишки человека HCT116 была выращена в среде McCoy’s 5A с добавлением 10% фетальной телячьей сыворотки и 1% антибиотика/антимикотика (Invitrogen, Гранд-Айленд, Нью-Йорк, США) и культивирована при 37°C во влажной атмосфере 5% CO 2 Клетки высевали в 96-луночные планшеты по 3750 клеток/лунку для анализа метаболизма MTS.
Неочищенный экстракт и воздействие 5-ФУ
Исходные растворы 10 мг мл -1 неочищенных экстрактов актиномицетов и положительного контроля 5-фторурацила (5-ФУ) при 8 мМ (Sigma) готовили в диметилсульфоксиде (ДМСО). Через 24 часа после посева клетки подвергали воздействию серийных разведений неочищенных экстрактов актиномицетов и 5-ФУ или контрольного носителя ДМСО в течение 72 часов. Все испытуемые соединения, 5-ФУ или ДМСО серийно разводили в четыре раза в культуральной среде.
Оценка цитотоксичности
Чтобы определить реакцию раковых клеток на химиотерапевтические препараты и другие соединения в целевых скринингах, а также изучить сигнальные пути рака толстой кишки, мы протестировали противораковую активность неочищенных экстрактов актиномицетов в клетках HCT116.Активность оценивали через 72 часа воздействия неочищенного экстракта на клетки, а также параллельно с положительным контролем и контролем с носителем. Противораковую активность оценивали с помощью CellTiter 96 ® Aqueous Non-Radioactive Cell Proliferation Assay (Promega, Мэдисон, Висконсин, США) с использованием 4-(4,5-диметилатиазол-2-ил)-5-(3-карбоксиметоксифенил)- 2-(4-сульфофенил), внутренняя соль (MTS), согласно инструкции производителя. Вкратце, это колориметрический метод определения количества жизнеспособных клеток в анализах пролиферации, цитотоксичности или химиочувствительности.Водный анализ CellTiter 96 ® состоит из растворов MTS и реагента связывания электронов, феназинметосульфата (PMS). Противораковую активность каждого неочищенного экстракта актиномицетов определяли по снижению клеточной редукции MTS в формазановый продукт, растворимый в культуральной среде ткани. Абсорбция формазанового продукта при 490 нм может быть измерена непосредственно в 96-луночных планшетах для анализа без дополнительной обработки, что коррелирует с количеством живых клеток в культуре.Вкратце, 20 мкл раствора MTS/PMS (19:1) добавляли непосредственно в каждую лунку 96-луночного планшета, содержащего 150 мкл каждого разведения неочищенного экстракта актиномицетов в культуральной среде, и количество формазанового продукта измеряли через 1 час после выдерживания. инкубация с использованием устройства для считывания микропланшетов Bio-Rad Model 680 (Bio-Rad Hercules, Калифорния, США) при 490 нм.
Вкратце, это колориметрический метод определения количества жизнеспособных клеток в анализах пролиферации, цитотоксичности или химиочувствительности.Водный анализ CellTiter 96 ® состоит из растворов MTS и реагента связывания электронов, феназинметосульфата (PMS). Противораковую активность каждого неочищенного экстракта актиномицетов определяли по снижению клеточной редукции MTS в формазановый продукт, растворимый в культуральной среде ткани. Абсорбция формазанового продукта при 490 нм может быть измерена непосредственно в 96-луночных планшетах для анализа без дополнительной обработки, что коррелирует с количеством живых клеток в культуре.Вкратце, 20 мкл раствора MTS/PMS (19:1) добавляли непосредственно в каждую лунку 96-луночного планшета, содержащего 150 мкл каждого разведения неочищенного экстракта актиномицетов в культуральной среде, и количество формазанового продукта измеряли через 1 час после выдерживания. инкубация с использованием устройства для считывания микропланшетов Bio-Rad Model 680 (Bio-Rad Hercules, Калифорния, США) при 490 нм. Значения максимальной половинной ингибирующей концентрации (IC 50 ) определяли с помощью GraphPad Prism (версия 5 программного обеспечения GraphPad, Сан-Диего, Калифорния, США).
Значения максимальной половинной ингибирующей концентрации (IC 50 ) определяли с помощью GraphPad Prism (версия 5 программного обеспечения GraphPad, Сан-Диего, Калифорния, США).
Антимикробный скрининг сырых экстрактов
Антимикробная активность была оценена для 400 неочищенных экстрактов путем проведения скрининга против двух наиболее важных устойчивых к антибиотикам микроорганизмов, вызывающих внутрибольничные инфекции, в частности, устойчивых к метициллину Staphylococcus aureus COL (MRSA; Gill et al., 2005) и устойчивых к ванкомицину (фенотип vanA) Enterococcus faecium EF82 (VREF; Mato et al., 1996), с использованием среды Brain Heart Infusion (BHI) (DIFCO Laboratories, Детройт, США, 1 л деионизированной воды). Скрининг проводили в 96-луночных планшетах; каждый неочищенный экстракт, предварительно сконцентрированный до 10 мг/мл -1 в ДМСО, добавляли к выращенной в логарифмической фазе культуре (OD 600 нм = 0,04-0,06) до конечной концентрации 2,5% (об. /об.). Все образцы были двукратно серийно разведены в пять раз, в результате чего конечные концентрации тестируемых соединений находились в диапазоне от 250 до 7.81 мкг мл -1 . Были испытаны дополнительные разведения, значения до 0,03 мкг мл -1 , если концентрация 7,81 мкг мл -1 показала ингибирующую активность. Через 18 ч инкубации при 37°С путем визуального осмотра и спектрофотометрического анализа определяли минимальные ингибирующие концентрации. Активные сырые экстракты повторно тестировали для подтверждения результата.
/об.). Все образцы были двукратно серийно разведены в пять раз, в результате чего конечные концентрации тестируемых соединений находились в диапазоне от 250 до 7.81 мкг мл -1 . Были испытаны дополнительные разведения, значения до 0,03 мкг мл -1 , если концентрация 7,81 мкг мл -1 показала ингибирующую активность. Через 18 ч инкубации при 37°С путем визуального осмотра и спектрофотометрического анализа определяли минимальные ингибирующие концентрации. Активные сырые экстракты повторно тестировали для подтверждения результата.
Результаты
Из 421 выделенного и секвенированного штамма 408 были идентифицированы как актиномицеты в соответствии с поиском BLASTN в базе данных GenBank.Восемь из этих последовательностей были сочтены химерами в процессе представления последовательностей NCBI и были исключены из нашего анализа. Остальные 400 последовательностей использовались для расчета разрежения и оценки разнообразия культивируемых актиномицетов на архипелаге Мадейра. В целом образцы, полученные с архипелага, показывают, что в его отложениях содержится по крайней мере 71 различная операционная таксономическая единица (OTU), охватывающая 24 рода актиномицетов (таблица 1). Как и в большинстве исследований культивирования, подавляющее большинство изолятов представляли OTU из рода Streptomyces .Было извлечено почти такое же количество OTU Micromonospora , но было выделено только 11 штаммов Actinomadura по сравнению с 90 из Micromonospora , что демонстрирует избыточность разнообразия штаммов из последнего рода. Большинство других идентифицированных родов были представлены только одной или двумя OTU, даже при большом числе выделенных штаммов (табл. 1). Вероятно, это связано с использованием консервативного порогового значения для определения отдельных OTU (идентичность последовательности 97%), каждая из которых может охватывать более одного вида.Например, род Salinispora представлен только одной OTU, хотя известно, что из собранного осадка были выделены два вида Salinispora ( Salinispora pacifica и S.
В целом образцы, полученные с архипелага, показывают, что в его отложениях содержится по крайней мере 71 различная операционная таксономическая единица (OTU), охватывающая 24 рода актиномицетов (таблица 1). Как и в большинстве исследований культивирования, подавляющее большинство изолятов представляли OTU из рода Streptomyces .Было извлечено почти такое же количество OTU Micromonospora , но было выделено только 11 штаммов Actinomadura по сравнению с 90 из Micromonospora , что демонстрирует избыточность разнообразия штаммов из последнего рода. Большинство других идентифицированных родов были представлены только одной или двумя OTU, даже при большом числе выделенных штаммов (табл. 1). Вероятно, это связано с использованием консервативного порогового значения для определения отдельных OTU (идентичность последовательности 97%), каждая из которых может охватывать более одного вида.Например, род Salinispora представлен только одной OTU, хотя известно, что из собранного осадка были выделены два вида Salinispora ( Salinispora pacifica и S. arenicola) . Тем не менее, даже при такой консервативной группировке расчеты разрежения культивируемого разнообразия ясно показывают, что асимптота богатства OTU (насыщение) не достигнута (рис. 1). Эти результаты свидетельствуют о том, что дальнейший отбор проб и использование большего количества методов выделения, вероятно, приведут к выделению дополнительных OTU культивируемых актиномицетов из отложений архипелага.Статистическая оценка культивируемого богатства с использованием Chao1 также предполагает, что из собранных образцов донных отложений можно выделить больше OTU актиномицетов, возможно, еще 118 OTU (рис. 1).
arenicola) . Тем не менее, даже при такой консервативной группировке расчеты разрежения культивируемого разнообразия ясно показывают, что асимптота богатства OTU (насыщение) не достигнута (рис. 1). Эти результаты свидетельствуют о том, что дальнейший отбор проб и использование большего количества методов выделения, вероятно, приведут к выделению дополнительных OTU культивируемых актиномицетов из отложений архипелага.Статистическая оценка культивируемого богатства с использованием Chao1 также предполагает, что из собранных образцов донных отложений можно выделить больше OTU актиномицетов, возможно, еще 118 OTU (рис. 1).
Таблица 1. Название родов и количество OTU и штаммов для каждого рода, выделенных из морских отложений архипелага Мадейра .
Рисунок 1. Кривые разрежения изолированных актиномицетов морского происхождения с архипелага Мадейра . Как наблюдаемые (sobs), так и оценочные (chao) кривые разнообразия показывают, что насыщение не было достигнуто в отношении разнообразия культивируемых актиномицетов морского происхождения.
Чтобы понять эволюционные отношения между выделенными OTU, было построено филогенетическое дерево (рис. 2). Дерево показывает, что большинство OTU очень похожи на ранее зарегистрированные последовательности актиномицетов, доступные в GenBank, хотя мы обнаружили OTU в родах Streptomyces , Stackebrandtia и A ctinomycetospora , которые филогенетически различны. Кроме того, когда потребность в морской воде для роста включена в дерево (звездочки на рис. 2), мы видим, что это не является исключительным признаком уже описанных морских облигатных родов (т.э., Salinispora ). Скорее, потребность в морской воде для роста является широко распространенной характеристикой среди изолятов из многих OTU, что позволяет предположить, что актиномицеты морского происхождения из разных родов адаптированы к морским условиям.
Рисунок 2. Филогенетическое дерево максимального правдоподобия гена 16s рРНК из 71 репрезентативной OTU актиномицетов с архипелага Мадейра и их ближайшего соседа в GenBank . Дерево было создано с использованием 1279 п.н. и 1000 бутстрепов. Цифры в скобках — это регистрационные номера GenBank, а номер PTM относится к внутреннему справочному номеру коллекции.Звездочками отмечены те ОТЕ, в которых наблюдались штаммы, нуждающиеся в морской воде для роста. Bacillus methanolicus использовали в качестве внешней группы.
Дерево было создано с использованием 1279 п.н. и 1000 бутстрепов. Цифры в скобках — это регистрационные номера GenBank, а номер PTM относится к внутреннему справочному номеру коллекции.Звездочками отмечены те ОТЕ, в которых наблюдались штаммы, нуждающиеся в морской воде для роста. Bacillus methanolicus использовали в качестве внешней группы.
Чтобы определить приоритеты будущих мест отбора проб для улучшения усилий по биоразведке, мы стремились определить места, в которых были обнаружены самые разнообразные группы актиномицетов. Для этого анализы UniFrac использовались для изучения популяций актиномицетов двумя способами: во-первых, мы решили взвесить влияние актиномицетов, происходящих с суши, сравнив разнообразие между прибрежными (≤ 100 м от берега) и прибрежными (≤ 100 м от берега) видами.прибрежных районах (> 100 м от берега), и, во-вторых, мы хотели изучить влияние различных характеристик отложений местообитаний на разнообразие изолированных актиномицетов. С помощью этих группировок были созданы дендограммы несхожести UniFrac (рис. 3). Результаты показывают, что прибрежные и морские образцы дают статистически значимые разные популяции актиномицетов, что указывает на то, что будущие сборы должны быть направлены на сбор образцов глубоких отложений вдали от берега, чтобы выявить штаммы актиномицетов, отличные от штаммов, обнаруженных в прибрежных районах.С другой стороны, когда анализы проводились на репрезентативных местообитаниях, 11 местообитаний образовали четыре разные клады (рис. 3). За исключением местообитаний «скалистый берег» и «песок», не было обнаружено существенных филогенетических различий между культивируемыми актиномицетами из разных типов отложений, что позволяет предположить, что тип отложений не является характеристикой, определяющей или влияющей на филогенетическое разнообразие актиномицетов, культивируемых нашими методами. Когда кривые разрежения были применены для анализа усилий по отбору проб для каждой среды обитания, мы заметили, что все среды обитания были либо недостаточно отобраны, либо что применяемая методология не позволяла восстановить большое разнообразие актиномицетов (дополнительный рисунок 1).
3). Результаты показывают, что прибрежные и морские образцы дают статистически значимые разные популяции актиномицетов, что указывает на то, что будущие сборы должны быть направлены на сбор образцов глубоких отложений вдали от берега, чтобы выявить штаммы актиномицетов, отличные от штаммов, обнаруженных в прибрежных районах.С другой стороны, когда анализы проводились на репрезентативных местообитаниях, 11 местообитаний образовали четыре разные клады (рис. 3). За исключением местообитаний «скалистый берег» и «песок», не было обнаружено существенных филогенетических различий между культивируемыми актиномицетами из разных типов отложений, что позволяет предположить, что тип отложений не является характеристикой, определяющей или влияющей на филогенетическое разнообразие актиномицетов, культивируемых нашими методами. Когда кривые разрежения были применены для анализа усилий по отбору проб для каждой среды обитания, мы заметили, что все среды обитания были либо недостаточно отобраны, либо что применяемая методология не позволяла восстановить большое разнообразие актиномицетов (дополнительный рисунок 1). Расчеты индекса разнообразия Шеннона показали, что, основываясь на наших стратегиях культивирования, самыми разнообразными местообитаниями были места с наибольшим количеством изолятов (таблица 2), что позволяет предположить, что в отложениях Мадейры дальнейшие методы изоляции могут привести к дополнительному культивируемому разнообразию актиномицетов. Дополнительным подтверждением этого результата является тот факт, что из 662 обработанных проб морских отложений только 198 вернули актиномицеты.
Расчеты индекса разнообразия Шеннона показали, что, основываясь на наших стратегиях культивирования, самыми разнообразными местообитаниями были места с наибольшим количеством изолятов (таблица 2), что позволяет предположить, что в отложениях Мадейры дальнейшие методы изоляции могут привести к дополнительному культивируемому разнообразию актиномицетов. Дополнительным подтверждением этого результата является тот факт, что из 662 обработанных проб морских отложений только 198 вернули актиномицеты.
Рис. 3. Дендограммы различий UniFrac по местам обитания (вверху) и по группе прибрежных и морских проб (внизу) .
Таблица 2. Разнообразие и распространение актиномицетов в различных местообитаниях, отобранных на архипелаге Мадейра .
Из-за нашего интереса к химическим возможностям этих микроорганизмов было проведено более глубокое исследование двух OTU, включающих клады MAR4 и MAR1 ( Salinispora ). Наши результаты показали шесть штаммов с архипелага, которые были широко распространены среди ранее описанных кладов разнообразия MAR4 (Gallagher et al. , 2013; Рисунок 4), с возможностью того, что некоторые из них могут считаться новыми филотипами MAR4. Между тем, филогенетический анализ штаммов Salinispora показал присутствие штаммов S. pacifica и штаммов S. arenicola в Атлантике. Среди них были обнаружены четыре новых филотипа S. pacifica (S, P, Q и R, рис. 5), что свидетельствует о том, что усилия по биоразведке по-прежнему играют важную роль в открытии новых филотипов химически важных штаммов морских актиномицетов.
, 2013; Рисунок 4), с возможностью того, что некоторые из них могут считаться новыми филотипами MAR4. Между тем, филогенетический анализ штаммов Salinispora показал присутствие штаммов S. pacifica и штаммов S. arenicola в Атлантике. Среди них были обнаружены четыре новых филотипа S. pacifica (S, P, Q и R, рис. 5), что свидетельствует о том, что усилия по биоразведке по-прежнему играют важную роль в открытии новых филотипов химически важных штаммов морских актиномицетов.
Рисунок 4. Филогенетическое дерево максимального правдоподобия с использованием 16s рРНК (1279 п.н. и 1000 бутстрапов) из штаммов актиномицетов Мадейры, принадлежащих к ранее описанной группе MAR4 . Для создания дерева использовали эталонные штаммы MAR4. Показаны номера PTM штаммов Мадейры, за которыми следует темный закрашенный кружок. Streptomyces rubreogriseus использовали в качестве внешней группы.
Рисунок 5. Филогенетическое дерево методом максимального правдоподобия с использованием 16s рРНК (1279 п. н. и 1000 бутстрапов) из штаммов актиномицетов Мадейры, принадлежащих к ранее описанному роду Salinispora .Для создания дерева использовали штамм Salinispora , номер ссылки . Виды Salinispora названы, а в скобках за ними следует их инвентарный номер GenBank. Показаны номера PTM штаммов Мадейры. Verrucosispora maris использовали в качестве внешней группы.
н. и 1000 бутстрапов) из штаммов актиномицетов Мадейры, принадлежащих к ранее описанному роду Salinispora .Для создания дерева использовали штамм Salinispora , номер ссылки . Виды Salinispora названы, а в скобках за ними следует их инвентарный номер GenBank. Показаны номера PTM штаммов Мадейры. Verrucosispora maris использовали в качестве внешней группы.
Чтобы получить представление о биоактивном потенциале актиномицетов морского происхождения с архипелага Мадейра, сырые экстракты всех 400 актиномицетов были проверены на противораковую активность в клеточной линии карциномы толстой кишки человека HCT116 и на антимикробную активность против метициллин-резистентного Staphylococcus aureus (MRSA) и устойчивых к ванкомицину возбудителей Enterococcus faecium (VREF).Из них удалось выделить три биологически активных рода актиномицетов: Streptomyces , Micromonospora, Salinispora (табл. 3, 4). В качестве первого взгляда на фармацевтическое применение этих штаммов была измерена биологическая активность неочищенных экстрактов. Биологическая активность варьировала от 0,65 до 59,05 мкг мл -1 для клеточных линий HCT116 и ≤ 0,03–250 мкг мл -1 для антимикробной активности MRSA и VREF. Наиболее биоактивные штаммы принадлежали к роду Streptomyces , а представители морской клады MAR4 были биоактивны во всех тестах.Изолят S. arenicola показал противоопухолевую активность (как и ожидалось) и антимикробную активность в отношении MRSA при 62 мкг/мл -1 (таблица 4). Удивительно, но ни один из изолятов S. pacifica не показал биологической активности ни в одном из биоанализов, что свидетельствует о том, что одни и те же виды, выделенные из разных мест в океане, могут различаться по профилям вторичных метаболитов или что методы культивирования необходимо оптимизировать для производства соединений. .
Биологическая активность варьировала от 0,65 до 59,05 мкг мл -1 для клеточных линий HCT116 и ≤ 0,03–250 мкг мл -1 для антимикробной активности MRSA и VREF. Наиболее биоактивные штаммы принадлежали к роду Streptomyces , а представители морской клады MAR4 были биоактивны во всех тестах.Изолят S. arenicola показал противоопухолевую активность (как и ожидалось) и антимикробную активность в отношении MRSA при 62 мкг/мл -1 (таблица 4). Удивительно, но ни один из изолятов S. pacifica не показал биологической активности ни в одном из биоанализов, что свидетельствует о том, что одни и те же виды, выделенные из разных мест в океане, могут различаться по профилям вторичных метаболитов или что методы культивирования необходимо оптимизировать для производства соединений. .
Таблица 3.Антимикробные результаты для неочищенных экстрактов морских актиномицетов против метициллин-резистентных Staphylococcus aureus COL (MRSA) и ванкомицин-резистентных (vanA фенотип) Enterococcus faecium EF82 (VREF) .
Таблица 4. Цитотоксическая активность (IC 50 мкг мл -1 ) неочищенных экстрактов морских актиномицетов в отношении клеток карциномы толстой кишки человека HCT116 в течение 72 ч .
Обсуждение
Португальский архипелаг Мадейра расположен в атлантическом регионе Макаронезии, который выходит из Африканской тектонической плиты и находится на южной оконечности хребта Торе-Мадейра.Регион имеет уникальную биогеографию и биоразнообразие. Поверхностные океанические течения Мадейры связаны с системой течений Северной Атлантики и не создаются ветрами Азорского антициклона. Глубоководная масса состоит из вод Северной Атлантики, Южной Атлантики и Средиземного моря (Emery, Meincke, 1986; Cox, 1989). Эти уникальные характеристики в сочетании с тем фактом, что нет никаких сообщений о культивируемых актиномицетах из отложений Атлантического океана, кроме Вейланда (Weyland, 1969), делают отложения архипелага Мадейра идеальным объектом для научных исследований в области биоразведки морских отложений.
Наше биоразведочное исследование нацелено на культивируемое разнообразие актиномицетов, к которому можно получить доступ с помощью недорогих и простых методов, включая пробоотборник, предназначенный для развертывания с небольшой лодки без необходимости платить за дорогостоящее время доставки.
Отложения оказались богатыми актиномицетами, в результате чего было обнаружено 400 культивируемых актиномицетов из 198 морских отложений. Всего из отложений, собранных в различных местах архипелага Мадейра, было культивировано 24 рода, в том числе 71 различный OTU.Кроме того, предполагаемые расчеты культивируемого разнообразия предполагают, что эти морские отложения обладают дополнительным потенциалом в качестве источника разнообразных актиномицетов. Большая часть культивируемого разнообразия принадлежала роду Streptomyces , где наличие 25 OTU показало, что этот род по-прежнему является одним из наиболее доступных для культивирования в морских отложениях, даже несмотря на то, что недавние исследования показали, что другие роды актиномицетов могут доминировать. популяции морских отложений (Gontang et al., 2007; Клавериас и др., 2015). Эти результаты, скорее всего, свидетельствуют о том, что наши методы культивирования были селективными для этого рода. Наши методы отбора проб действительно привели к выделению 24 различных родов, что значительно превышает разнообразие актиномицетов, культивируемых из атлантических морских отложений на сегодняшний день (Weyland, 1969, 1984; Stach et al., 2003; Maldonado et al., 2005; Duncan et al., 2014, 2015). Наше пороговое значение 97% для группировки OTU привело к консервативному количеству OTU, даже когда количество выделенных штаммов было высоким (например,г., Salinispora, Nocardiopsis) . Использование консервативного порогового значения не предназначалось для определения видов бактерий. Таким образом, мы предполагаем, что наши 400 изолятов представляют более 71 вида бактерий. Поскольку эти OTU имеют > 98% идентичности последовательности гена 16S рРНК со штаммами, о которых уже сообщалось в общедоступных базах данных, мы не считаем какие-либо из изолятов новыми штаммами.
популяции морских отложений (Gontang et al., 2007; Клавериас и др., 2015). Эти результаты, скорее всего, свидетельствуют о том, что наши методы культивирования были селективными для этого рода. Наши методы отбора проб действительно привели к выделению 24 различных родов, что значительно превышает разнообразие актиномицетов, культивируемых из атлантических морских отложений на сегодняшний день (Weyland, 1969, 1984; Stach et al., 2003; Maldonado et al., 2005; Duncan et al., 2014, 2015). Наше пороговое значение 97% для группировки OTU привело к консервативному количеству OTU, даже когда количество выделенных штаммов было высоким (например,г., Salinispora, Nocardiopsis) . Использование консервативного порогового значения не предназначалось для определения видов бактерий. Таким образом, мы предполагаем, что наши 400 изолятов представляют более 71 вида бактерий. Поскольку эти OTU имеют > 98% идентичности последовательности гена 16S рРНК со штаммами, о которых уже сообщалось в общедоступных базах данных, мы не считаем какие-либо из изолятов новыми штаммами. Тем не менее потребность в морской воде для роста предполагает, что некоторые из изолятов могут быть новыми видами или новыми экотипами.Имеются данные о том, что огромное культивируемое разнообразие актиномицетов, обнаруженное в отложениях Мадейры, может быть расширено, поскольку насыщение OTU не было достигнуто, а оценщик разнообразия Chao1 предполагает, что из этих отложений можно выделить еще 118 OTU (рис. 1). В будущем следует расширить усилия по культивированию и использовать больше сред или методов предварительной обработки для доступа к дополнительному разнообразию (Janssen et al., 2002). Адаптации к морской среде, о чем свидетельствует потребность в морской воде для роста, можно наблюдать у большинства родов, выделенных в этом исследовании.Хотя было описано лишь несколько морских облигатных актиномицетов (Maldonado et al., 2005; Tian et al., 2012), большое количество штаммов, нуждающихся в морской воде, позволяет предположить, что архипелаг Мадейра является источником актиномицетов, адаптированных к жизни в морской среде.
Тем не менее потребность в морской воде для роста предполагает, что некоторые из изолятов могут быть новыми видами или новыми экотипами.Имеются данные о том, что огромное культивируемое разнообразие актиномицетов, обнаруженное в отложениях Мадейры, может быть расширено, поскольку насыщение OTU не было достигнуто, а оценщик разнообразия Chao1 предполагает, что из этих отложений можно выделить еще 118 OTU (рис. 1). В будущем следует расширить усилия по культивированию и использовать больше сред или методов предварительной обработки для доступа к дополнительному разнообразию (Janssen et al., 2002). Адаптации к морской среде, о чем свидетельствует потребность в морской воде для роста, можно наблюдать у большинства родов, выделенных в этом исследовании.Хотя было описано лишь несколько морских облигатных актиномицетов (Maldonado et al., 2005; Tian et al., 2012), большое количество штаммов, нуждающихся в морской воде, позволяет предположить, что архипелаг Мадейра является источником актиномицетов, адаптированных к жизни в морской среде. океан.
океан.
Статистическая разница между культивируемым разнообразием актиномицетов, наблюдаемым в отложениях, близких к берегу, и в прибрежных отложениях, подчеркивает важность поездок для сбора, которые включают последние, удаленные места. Кроме того, отчетливое культивируемое филогенетическое разнообразие было обнаружено в местообитаниях, состоящих из камней и песка, хотя ни одно другое местообитание из оставшихся девяти, из которых были отобраны образцы, не указывало на сильную связь между типом отложений и присутствующим филогенетическим разнообразием культивируемых актиномицетов.Это наблюдение было также зарегистрировано Дунканом и его сотрудниками, которые исследовали распространение актиномицетов в морских отложениях из Нью-Брансуика, Канада (Duncan et al., 2014). Из 11 исследованных местообитаний «глубокий песчаный край каменистого склона» обеспечил наиболее разнообразную совокупность изолятов (таблица 2), и это более высокое относительное культивируемое разнообразие может быть связано с тем фактом, что в среднем на образец было выделено три штамма. в то время как из других местообитаний восстановление штаммов было ниже двух на образец.
в то время как из других местообитаний восстановление штаммов было ниже двух на образец.
Хотя наше исследование было направлено на культивируемое разнообразие актиномицетов, мы считаем, что общее богатство актиномицетов в этих отложениях выше представленного здесь. Это можно предположить из двух источников информации. Во-первых, наша кривая разрежения не достигла насыщения (рис. 1), что свидетельствует о наличии более культивируемых штаммов, а во-вторых, недавние независимые от культивирования исследования морских отложений показали, что 13% разнообразия в них принадлежит классу Actinobacteria. (Бинхольд и др., 2016). Хотя легкость доступа к этому разнообразию не установлена, внутренние характеристики отложений можно использовать в качестве ориентира для разработки новых методов культивирования и внесения этих актиномицетов в лабораторную культуру. Исследования, подобные представленному здесь, показывают, что обширность океанических морских отложений позволяет легко применять методы отбора проб и культивирования для доступа к широкому разнообразию актиномоцитов морского происхождения с потенциалом обнаружения природных продуктов.
Одной из наиболее важных особенностей биоразведки, помимо открытия новой культивируемой филогении, является определение биоактивного потенциала выделенных штаммов.Соединяя эти два компонента вместе, можно выделить штаммы, принадлежащие к разным морским группам новой филогении, и оценить их способность производить интересный химический состав. Так обстоит дело с актиномицетами, принадлежащими к морской кладе MAR4, поскольку сообщалось, что они являются важным источником гибридных изопреноидов (Gallagher et al., 2010), и была предложена связь между их филогении и соединениями, которые они продуцируют (Gallagher et al., 2010). и др., 2013). В этом исследовании были выделены различные штаммы MAR4, и некоторые из них показали интересную биологическую активность (см. Таблицы 3, 4).Хотя ни один из филотипов S. pacifica не обладал биологической активностью (по сравнению с одним филотипом S. arenicola , обладающим как противомикробной, так и цитотоксической активностью), их способность производить новый химический состав и проявлять различную биологическую активность еще предстоит изучить (Freel et al. , 2011; Vidgen et al., 2012; Bose et al., 2014). Кроме того, наши усилия по отбору образцов привели к первому сообщению о Salinispora spp. из умеренных вод Северной Атлантики. Salinispora spp.ранее были выделены только из более теплых тропических вод (Jensen et al., 2015).
, 2011; Vidgen et al., 2012; Bose et al., 2014). Кроме того, наши усилия по отбору образцов привели к первому сообщению о Salinispora spp. из умеренных вод Северной Атлантики. Salinispora spp.ранее были выделены только из более теплых тропических вод (Jensen et al., 2015).
Был оценен биоактивный потенциал 400 штаммов, и было обнаружено, что 22 штамма имеют активные неочищенные экстракты либо как противомикробные, либо как цитотоксические агенты. Интересно отметить, что, несмотря на то, что в нашей коллекции было в общей сложности 24 идентифицируемых рода, только три показали биологическую активность в наших анализах, что позволяет предположить, что изучение морских Streptomyces, Micromonopora и Salinispora по-прежнему является важным и плодотворный в отношении открытия новых натуральных продуктов.
В морских отложениях архипелага Мадейра обнаружено разнообразное сообщество морских актиномицетов, самое большое разнообразие, когда-либо наблюдаемое в Атлантике. Наше биоразведочное исследование показало, что различные популяции этих микроорганизмов могут быть доступны как в прибрежных, так и в морских районах. Учитывая способность морских актиномицетов производить биологически активные соединения, которая была показана как в настоящем, так и в предыдущих исследованиях, мы предлагаем предпринять усилия для получения доступа к морским популяциям актиномицетов и использовать более интенсивные методы культивирования для доступа к огромному разнообразию потенциально плодовитых видов. производители химических веществ в океане.
Наше биоразведочное исследование показало, что различные популяции этих микроорганизмов могут быть доступны как в прибрежных, так и в морских районах. Учитывая способность морских актиномицетов производить биологически активные соединения, которая была показана как в настоящем, так и в предыдущих исследованиях, мы предлагаем предпринять усилия для получения доступа к морским популяциям актиномицетов и использовать более интенсивные методы культивирования для доступа к огромному разнообразию потенциально плодовитых видов. производители химических веществ в океане.
Вклад авторов
Все перечисленные авторы внесли существенный, непосредственный и интеллектуальный вклад в работу и одобрили ее для публикации.
Финансирование
Эта работа была поддержана финансированием от FCT/MEC через гранты PTDC/QUIQUI/119116/2010, IF/00700/2014, UID/QUI/50006/2013 (LAQV) и UID/Multi/04378/2013 (UCIBIO) и софинансируется ЕФРР в рамках соглашения о партнерстве PT2020 POCI-01-0145-FEDER — 007265 и POCI-01-0145-FEDER-007728. Финансирование 7-й рамочной программы ЕС (FP7/2007–2013) в рамках соглашения о гранте PCOFUND-GA-2009-246542 и 269138-NanoGuard. YP признает CONACYT для получения степени магистра наук. стипендия 560614.
Финансирование 7-й рамочной программы ЕС (FP7/2007–2013) в рамках соглашения о гранте PCOFUND-GA-2009-246542 и 269138-NanoGuard. YP признает CONACYT для получения степени магистра наук. стипендия 560614.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Рецензент SLL и редактор-обработчик заявили о своей общей принадлежности, а редактор-обработчик заявляет, что процесс, тем не менее, соответствует стандартам справедливого и объективного обзора.
Благодарности
Авторы выражают признательность У. Феникалу, П. Р. Дженсену и К. А. Кауфману из Океанографического института Скриппса, Сан-Диего, США, за поддержку, предоставленную для сбора проб, включая обращение с устройством для отбора проб отложений. П. Р. Дженсен, Н. Миллан и К. Галлахер получили признание за научные дискуссии о филогении 1 марта и 4 марта. Мы благодарим Э. Гонтанг за ее ценную помощь в написании этой статьи. Мы признаем П.Castilho из Университета Мадейры и M. Freitas из Морской биологической станции Фуншала за их гостеприимство и материально-техническую поддержку во время полевого сбора. UCIBIO и LAQV, REQUIMTE также благодарны за исследовательское оборудование. Мы признательны за любезный дар штаммов, использованных в этой работе: штамм MRSA COL был предоставлен H. de Lencastre (ITQB/UNL, Португалия), а штамм VREF EF82 – R. Mato (FCT/UNL, Португалия).
Мы благодарим Э. Гонтанг за ее ценную помощь в написании этой статьи. Мы признаем П.Castilho из Университета Мадейры и M. Freitas из Морской биологической станции Фуншала за их гостеприимство и материально-техническую поддержку во время полевого сбора. UCIBIO и LAQV, REQUIMTE также благодарны за исследовательское оборудование. Мы признательны за любезный дар штаммов, использованных в этой работе: штамм MRSA COL был предоставлен H. de Lencastre (ITQB/UNL, Португалия), а штамм VREF EF82 – R. Mato (FCT/UNL, Португалия).
Дополнительный материал
Дополнительный материал к этой статье можно найти в Интернете по адресу: https://www.frontiersin.org/article/10.3389/fmicb.2016.01594
Ссылки
Асолкар Р. Н., Фрил К. С., Дженсен П. Р., Феникал В., Кондратюк Т. П., Парк Э.-Дж. и др. (2009). Аренамиды, A.-C., цитотоксические ингибиторы NF каппа B из морского актиномицета Salinispora arenicola . J. Nat. Произв. 72, 396–402. дои: 10.1021/np800617a
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Бесеррил-Эспиноса, А. , Фрил, К.С., Дженсен, П.Р. и Сориа-Меркадо, И. Э. (2013). Морские Actinobacteria из Калифорнийского залива: разнообразие, численность и потенциал биосинтеза вторичных метаболитов. Антони Ван Левенгук 103, 809–819. doi: 10.1007/s10482-012-9863-3
, Фрил, К.С., Дженсен, П.Р. и Сориа-Меркадо, И. Э. (2013). Морские Actinobacteria из Калифорнийского залива: разнообразие, численность и потенциал биосинтеза вторичных метаболитов. Антони Ван Левенгук 103, 809–819. doi: 10.1007/s10482-012-9863-3
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Бинхольд, К., Зингер, Л., Боэтиус, А., и Раметт, А. (2016). Разнообразие и биогеография батиальных и абиссальных донных бактерий. PLoS ONE 11:e0148016.doi: 10.1371/journal.pone.0148016
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Bose, U., Hewavitharana, A.K., Vidgen, M.E., Ng, Y.K., Shaw, P.N., Fuerst, J.A., et al. (2014). Обнаружение сложного спектра вторичного метаболома видов Salinispora : исследование межвидового разнообразия. PLoS ONE 9:e91488. doi: 10.1371/journal.pone.0091488
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Бредхолт, Х. , Fjaervik, E., Johnsen, G., and Zotchev, S.B. (2008). Актиномицеты из отложений Тронхеймского фьорда, Норвегия: разнообразие и биологическая активность. Мар. Наркотики 6, 12–24. дои: 10.3390/md6010012
, Fjaervik, E., Johnsen, G., and Zotchev, S.B. (2008). Актиномицеты из отложений Тронхеймского фьорда, Норвегия: разнообразие и биологическая активность. Мар. Наркотики 6, 12–24. дои: 10.3390/md6010012
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Клавериас Ф.П., Ундабаррена А., Гонсалес М., Сигер М. и Камара Б. (2015). Культивируемое разнообразие и антимикробная активность Actinobacteria из морских отложений в заливе Вальпараисо, Чили. Перед. микробиол. 6:737. doi: 10.3389/fmicb.2015.00737
Реферат PubMed | Полный текст перекрестной ссылки
Кокс, доктор медицины (1989). Идеализированная модель мирового океана 1. Водные массы глобального масштаба. J. Phys. океаногр. 19, 1730–1752 гг.
Академия Google
Дхармарадж, С. (2010). Морской Streptomyces как новый источник биоактивных веществ. World J. Microbiol. Биотехнолог. 26, 2123–2139. doi: 10.1007/s11274-010-0415-6
Полнотекстовая перекрестная ссылка | Академия Google
Дункан, К. , Haltli, B., Gill, K.A., and Kerr, R.G. (2014). Биоразведка морских отложений Нью-Брансуика, Канада: изучение взаимосвязи между общим бактериальным разнообразием и разнообразием Actinobacteria . Мар. Наркотики 12, 899–925. дои: 10.3390/md12020899
, Haltli, B., Gill, K.A., and Kerr, R.G. (2014). Биоразведка морских отложений Нью-Брансуика, Канада: изучение взаимосвязи между общим бактериальным разнообразием и разнообразием Actinobacteria . Мар. Наркотики 12, 899–925. дои: 10.3390/md12020899
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Дункан, К. Р., Халтли, Б., Гилл, К. А., Корреа, Х., Берру, Ф., и Керр, Р. Г. (2015). Изучение разнообразия и метаболического потенциала актиномицетов из умеренных морских отложений Ньюфаундленда, Канада. J. Ind. Microbiol. Биотехнолог. 42, 57–72. doi: 10.1007/s10295-014-1529-x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Эмери, В. Дж., и Мейнке, Дж. (1986). Глобальные водные массы — резюме и обзор. Океанол. Acta 9, 383–391.
Академия Google
Fiedler, H.P., Bruntner, C., Bull, A.T., Ward, A.C., Goodfellow, M., Potterat, O., et al. (2005). Морские актиномицеты как источник новых вторичных метаболитов. Антони Ван Левенгук 87, 37–42. doi: 10.1007/s10482-004-6538-8
Антони Ван Левенгук 87, 37–42. doi: 10.1007/s10482-004-6538-8
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Freel, K.C., Nam, S.-J., Fenical, W., and Jensen, P.R. (2011). Эволюция генов вторичных метаболитов у трех близкородственных морских видов актиномицетов. Заяв. Окружающая среда. микробиол. 77, 7261–7270. doi: 10.1128/AEM.05943-11
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Галлахер, К.А., Феникал В. и Дженсен П.Р. (2010). Гибридная продукция вторичных метаболитов изопреноидов у наземных и морских актиномицетов. Курс. мнение Биотехнолог. 21, 794–800. doi: 10.1016/j.copbio.2010.09.010
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Галлахер, К. А., Раушер, К., Иока, Л. П., и Дженсен, П. Р. (2013). Филогенетическое и химическое разнообразие гибридно-изопреноидпродуцирующей линии Streptomycete . Заяв. Окружающая среда.микробиол. 79, 6894–6902. doi: 10.1128/AEM.01814-13
79, 6894–6902. doi: 10.1128/AEM.01814-13
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Gill, S.R., Fouts, D.E., Archer, G.L., Mongodin, E.F., DeBoy, R.T., Ravel, J., et al. (2005). Представления об эволюции вирулентности и резистентности на основе полного анализа генома раннего метициллин-резистентного штамма Staphylococcus aureus и биопленкообразующего метициллин-резистентного штамма Staphylococcus epidermidis . Дж.бактериол. 187, 2426–2438. doi: 10.1128/JB.187.7.2426-2438.2005
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Гонтанг, Э.А., Феникал, В., и Дженсен, П.Р. (2007). Филогенетическое разнообразие грамположительных бактерий, культивируемых из морских отложений. Заяв. Окружающая среда. микробиол. 73, 3272–3282. doi: 10.1128/AEM.02811-06
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Янссен, П. Х., Йейтс, П. С., Гринтон, Б. Э., Тейлор, П.М., и Саит, М. (2002). Улучшенная культивируемость почвенных бактерий и выделение в чистой культуре новых членов отделов Acidobacteria, Actinobacteria, Proteobacteria и Verrucomicrobia . Заяв. Окружающая среда. микробиол. 68, 2391–2396. doi: 10.1128/AEM.68.5.2391-2396.2002
Э., Тейлор, П.М., и Саит, М. (2002). Улучшенная культивируемость почвенных бактерий и выделение в чистой культуре новых членов отделов Acidobacteria, Actinobacteria, Proteobacteria и Verrucomicrobia . Заяв. Окружающая среда. микробиол. 68, 2391–2396. doi: 10.1128/AEM.68.5.2391-2396.2002
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Дженсен П.Р., Гонтанг Э., Мафнас К., Минсер Т.Дж. и Феникал В. (2005b). Культивируемое разнообразие морских актиномицетов из тропических отложений Тихого океана. Окружающая среда. микробиол. 7, 1039–1048. doi: 10.1007/s10482-004-6540-1
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Дженсен, П. Р., Минсер, Т. Дж., Уильямс, П. Г., и Феникал, В. (2005a). Разнообразие морских актиномицетов и открытие натуральных продуктов. Антони Ван Левенгук 87, 43–48. doi: 10.1007/s10482-004-6540-1
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Дженсен, П. Р., Мур, Б. С., и Феникал, В.(2015). Род морских актиномицетов Salinispora : модельный организм для обнаружения вторичных метаболитов. Нац. Произв. 32, 738–751. дои: 10.1039/c4np00167b
Р., Мур, Б. С., и Феникал, В.(2015). Род морских актиномицетов Salinispora : модельный организм для обнаружения вторичных метаболитов. Нац. Произв. 32, 738–751. дои: 10.1039/c4np00167b
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Лозупоне, К., и Найт, Р. (2005). UniFrac: новый филогенетический метод сравнения микробных сообществ. Заяв. Окружающая среда. микробиол. 71, 8228–8235. doi: 10.1128/AEM.71.12.8228-8235.2005
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Магарви, Н.А., Келлер, Дж. М., Бернан, В., Дворкин, М., и Шерман, Д. Х. (2004). Выделение и характеристика новых таксонов актиномицетов морского происхождения, богатых биоактивными метаболитами. Заяв. Окружающая среда. микробиол. 70, 7520–7529. doi: 10.1128/AEM.70.12.7520-7529.2004
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Мальдонадо, Л. А., Феникал, В., Дженсен, П.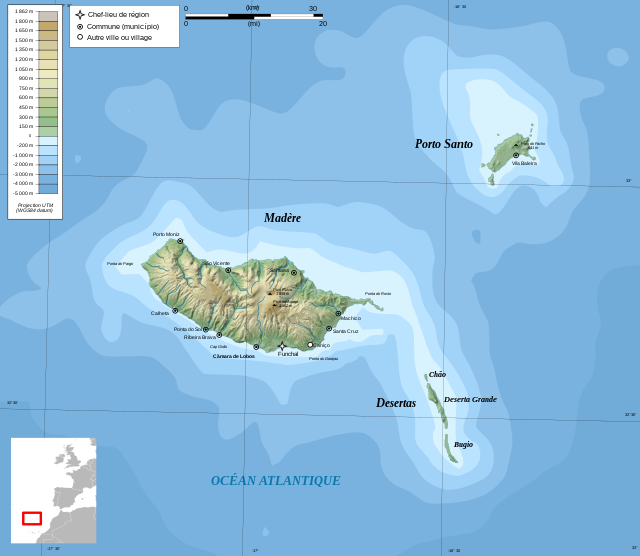 Р., Кауфман, К. А., Минсер, Т. Дж., Уорд, А. С., и соавт. (2005). Salinispora arenicola род. ноябрь, sp nov и Salinispora tropica sp nov., облигатные морские актиномицеты, принадлежащие к семейству Micromonosporaceae . Междунар. Дж. Сист. Эвол. микробиол. 55, 1759–1766. doi: 10.1099/ijs.0.63625-0
Р., Кауфман, К. А., Минсер, Т. Дж., Уорд, А. С., и соавт. (2005). Salinispora arenicola род. ноябрь, sp nov и Salinispora tropica sp nov., облигатные морские актиномицеты, принадлежащие к семейству Micromonosporaceae . Междунар. Дж. Сист. Эвол. микробиол. 55, 1759–1766. doi: 10.1099/ijs.0.63625-0
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Мартини А.К., Тай А.П.К., Венециано Д., Примо Ф. и Чисхолм С.В. (2009). Таксономическое разрешение, экотипы и биогеография Prochlorococcus . Окружающая среда.микробиол. 11, 823–832. doi: 10.1111/j.1462-2920.2008.01803.x.
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Мато Р., де Ленкастр Х., Робертс Р. Б. и Томаш А. (1996). Множественность генетического фона среди устойчивых к ванкомицину изолятов Enterococcus faecium , извлеченных из вспышки в больнице Нью-Йорка. Микроб. Сопротивление наркотикам. 2, 309–317. doi: 10.1089/mdr.1996.2.309
doi: 10.1089/mdr.1996.2.309
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Немергут, д.R., Costello, E.K., Hamady, M., Lozupone, C., Jiang, L., Schmidt, S.K., et al. (2011). Глобальные закономерности в биогеографии бактериальных таксонов. Окружающая среда. микробиол. 13, 135–144. doi: 10.1111/j.1462-2920.2010.02315.x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Патом-Ари, В., Стах, Дж. Э. М., Уорд, А. С., Хорикоши, К., Булл, А. Т., и Гудфеллоу, М. (2006). Разнообразие актиномицетов, выделенных из отложений Бездны Челленджера (10 898 м) Марианской впадины. Экстремофилы 10, 181–189. doi: 10.1007/s00792-005-0482-z
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Патин, Н. В., Дункан, К. Р., Доррестейн, П. К., и Дженсен, П. Р. (2016). Конкурентные стратегии позволяют дифференцировать близкородственные виды морских актинобактерий . ISME J. 10, 478–490. doi: 10.1038/ismej.2015.128
10, 478–490. doi: 10.1038/ismej.2015.128
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Пенн, К., Дженкинс, К., Нетт, М., Udwary, D.W., Gontang, E.A., McGlinchey, R.P., et al. (2009). Геномные островки связывают вторичный метаболизм с функциональной адаптацией у морских актинобактерий . ISME J. 3, 1193–1203. doi: 10.1038/ismej.2009.58
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Прието-Даво, А., Вильярреал-Гомес, Л. Дж., Форшнер-Данкос, С., Булл, А. Т., Стах, Дж. Э., Смит, Д. К., и соавт. (2013). Целевой поиск актиномицетов из прибрежных и глубоководных морских отложений. FEMS микробиол. Экол. 84, 510–518. дои: 10.1111/1574-6941.12082
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Prudhomme, J., McDaniel, E., Ponts, N., Bertani, S., Fenical, W., Jensen, P., et al. (2008). Морской Actinomycetes : новый источник соединений против малярийного паразита человека. PLoS ONE 3:e2335. doi: 10.1371/journal.pone.0002335
PLoS ONE 3:e2335. doi: 10.1371/journal.pone.0002335
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Рахман, М.А., Ислам, М.З., и Ислам, М.А.У. (2011). Антибактериальная активность изолятов актиномицетов, собранных из почв Раджшахи, Бангладеш. Биотехнолог. Рез. Междунар. 2011, 857925–857925. дои: 10.4061/2011/857925
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Schloss, P.D., Westcott, S.L., Ryabin, T., Hall, J.R., Hartmann, M., Hollister, E.B., et al. (2009). Представляем mothur: открытое, независимое от платформы, поддерживаемое сообществом программное обеспечение для описания и сравнения микробных сообществ. Заяв. Окружающая среда. микробиол. 75, 7537–7541. doi: 10.1128/AEM.01541-09
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Солано Г., Рохас-Хименес К., Джаспарс М. и Тамайо-Кастильо Г. (2009). Изучение разнообразия культивируемых актиномицетов на северном тихоокеанском и карибском побережьях Коста-Рики. Антони Ван Левенгук 96, 71–78. doi: 10.1007/s10482-009-9337-4
Антони Ван Левенгук 96, 71–78. doi: 10.1007/s10482-009-9337-4
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Стах, Дж.Э. М., Мальдонадо, Л. А., Массон, Д. Г., Уорд, А. С., Гудфеллоу, М., и Булл, А. Т. (2003). Статистические подходы к оценке разнообразия актинобактерий в морских отложениях. Заяв. Окружающая среда. микробиол. 69, 6189–6200. doi: 10.1128/AEM.69.10.6189-6200.2003
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Такидзава М., Колвелл Р. Р. и Хилл Р. Т. (1993). Выделение и разнообразие актиномицетов в Чесапикском заливе. Заяв. Окружающая среда. микробиол. 59, 997–1002.
Реферат PubMed | Академия Google
Тамура К., Стечер Г., Петерсон Д., Филипски А. и Кумар С. (2013). MEGA6: молекулярно-эволюционный генетический анализ версии 6.0. Мол. биол. Эвол. 30, 2725–2729. doi: 10.1093/molbev/mst197
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Tian, X. -P., Xu, Y., Zhang, J., Li, J., Chen, Z., Kim, C.-J., et al. (2012). Streptomyces oceani sp nov., новый облигатный морской актиномицет, выделенный из глубоководного образца просачивающейся аутигенной карбонатной конкреции в Южно-Китайском море. Антони Ван Левенгук 102, 335–343. doi: 10.1007/s10482-012-9743-x
-P., Xu, Y., Zhang, J., Li, J., Chen, Z., Kim, C.-J., et al. (2012). Streptomyces oceani sp nov., новый облигатный морской актиномицет, выделенный из глубоководного образца просачивающейся аутигенной карбонатной конкреции в Южно-Китайском море. Антони Ван Левенгук 102, 335–343. doi: 10.1007/s10482-012-9743-x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Видген, М.Е., Хупер, Дж.Н.А., и Фюрст, Дж.А. (2012). Разнообразие и распространение биоактивных актинобактерий рода Salinispora из губок Большого Барьерного рифа. Антони Ван Левенгук 101, 603–618. doi: 10.1007/s10482-011-9676-9
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Уокер, Дж.Д. и Колвелл Р. Р. (1975). Факторы, влияющие на подсчет и изоляцию актиномицетов из отложений Чесапикского залива и юго-восточной части Атлантического океана. Мар. Биол. 30, 193–201.
Академия Google
Вейланд, Х. (1984). «Актиномицеты донных отложений различных морей», в 2 Deuxième Colloque International de Bactériologie Marine — CNRS (Брест: IFREMER), 73–79.
(1984). «Актиномицеты донных отложений различных морей», в 2 Deuxième Colloque International de Bactériologie Marine — CNRS (Брест: IFREMER), 73–79.
Уильямс, Д. Э., Бернан, В. С., Ритакко, Ф. В., Мейзе, В. М., Гринштейн, М.и Андерсен, Р. Дж. (1999). Holyrines A и B, возможные промежуточные продукты биосинтеза стауроспорина, продуцируемые в культуре морскими актиномицетами, полученными из северной части Атлантического океана. Тетраэдр Летт. 40, 7171–7174. doi: 10.1016/S0040-4039(99)01495-1
Полнотекстовая перекрестная ссылка | Академия Google
Файл:Архипелаг Мадейры.png — Викисклад
| ОписаниеМадейра архипелаг.png |
Карта ареала Regulus madeirensis на основе Малларни, Киллиана; Свенссон, Ларс; Зеттерстрем, Дэн; Грант, Питер (1999). Путеводитель по птицам Коллинза . Лондон: Коллинз. п. 336. ISBN 000219728-6. Deutsch: Positionskarte von Madeira, Portugal Географическая карта: * N: 33,16° N * S: 32,38° N * W: 17,31° W * O: 16,23° W English: Карта расположения Мадейры, Португалия Географические границы карты: * N: 33,16° N * S: 32,38° N * W: 17,31° W * E: 16,23° W |
| Дата |
5 июля 2010 г. , 07:49 (UTC) , 07:49 (UTC) |
| Источник | |
| Автор |
|
|
Этот файл находится под лицензией Creative Commons Attribution-Share Alike 3.0 Непортированная лицензия. | |
https://creativecommons.org/licenses/by-sa/3.0 CC BY-SA 3.0 Creative Commons Attribution-Share Alike 3.0 правда правда |

Исходный журнал загрузки[править]
Это изображение является производным от следующих изображений:
- Файл:Portugal_Madeira_location_map.svg с лицензией Cc-by-sa-2.0
- 2009-12-19T15:41:01Z NordNordWest 1590×1365 (105588 Bytes) == {{int:filedesc}} == {{Information |Description= {{de|Positionskarte von [[:de:Madeira|Madeira] ], [[:de:Португалия|Португалия]]}} Географическая карта: * N: 33,16° N * S: 32,38° N * W: 17,31° W * O: 16,23° W {
Загружено с производной FX
Щелкните дату/время, чтобы просмотреть файл в том виде, в котором он был в то время.
| Дата / время | Дата | Размеры | Размеры | Комментарий | Комментарий |
|---|---|---|---|---|---|
07:52, 5 июля 2010 г. | 07:52, 5 июля 2010 г. 1000 000 × 858 (55 кб) | Jimfblak talk | contribs) | {{Information |Description={{de|Positionskarte von Madeira, Portugal}} Geographische Begrenzung der Karte: * N: 33,16° N * S: 32,38° N * W: 17,31° W * O: 16,23 ° W {{ru|Карта расположения Мадейры, [[:ru:Por |
Этот файл нельзя перезаписать.
Этот файл используется на следующей странице:
Этот файл используют следующие другие вики:
- Использование на an.wikipedia.org
- Использование на arz.wikipedia.org
- Использование на ast.wikipedia.org
- Использование на bg.wikipedia.org
- Использование на ca.wikipedia.org
- Использование на ceb.wikipedia.org
- Использование на en.wikipedia.org
- Использование на es.wikipedia.org
- Использование на eu.wikipedia.org
- Использование на fa.wikipedia.org
- Использование на it.
 wikipedia.org
wikipedia.org
- Использование на ru.wikipedia.org
- Использование на sk.wikipedia.org
- Использование на sv.wikipedia.org
- Использование на uk.wikipedia.org
- Использование на vi.wikipedia.org
- Использование на www.wikidata.org
Сколько островов на архипелаге Мадейра?
Обычно многие посетители острова, даже те, кто видел здесь давно, не знают, сколько островов на архипелаге Мадейра.
Архипелаг Мадейра расположен в Атлантическом океане между 30° и 33° северной широты, в 978 км к юго-западу от Лиссабона и примерно в 700 км от побережья Африки, почти на той же широте, что и Касабланка, относительно недалеко от Гибралтарского пролива.
Хотя обычно считается европейской территорией, географически архипелаг уже расположен на континентальном шельфе Африки. Ближайшим морским соседом является испанский архипелаг Канарские острова.
Сколько островов в архипелаге Мадейра?
1.
 Остров Мадейра
Остров Мадейра Острова архипелага Мадейра: Остров Мадейра – Изображение 2
Остров Мадейра – главный остров архипелага Мадейра, расположенный в Атлантическом океане, к юго-западу от португальского побережья, присоединенный к Европейскому Союз. Это, вместе с Порту-Санту, необитаемыми островами и островами Сельвагенс, архипелаг Мадейра и автономный регион Мадейра со столицей в городе Фуншал.
Остров Мадейра имеет вулканическое происхождение, площадь 742,4 км², обширную эндемическую флору, местную и экзотическую, а его типичный средиземноморский климат преобладает на южной стороне, переходящей в умеренный климат при более высоких квотах. Экономика широко ориентирована на туризм.
2. Остров Порту-Санту
Острова архипелага Мадейра: Остров Порту-Санту – Изображение 3
Порту-Санту – португальский муниципалитет, занимающий весь остров Порту-Санту, автономный район Мадейры, расположенный в деревне с другим название от муниципалитета, город Вила Балейра. Его площадь составляет 42,48 км².
Его площадь составляет 42,48 км².
Это один из шести муниципалитетов Португалии, в которых есть один приход с таким же названием. Ближайший муниципалитет — Машику на острове Мадейра, расположенный к юго-западу. Мадейры часто называют его Золотым островом.
3. Необитаемые острова
Острова архипелага Мадейра: Необитаемые острова – Изображение 4
Острова Дезерташ – это субархипелаг архипелага Мадейра, Португалия, вулканического происхождения, расположенный к юго-востоку от острова Мадейра.Это природный заповедник «Необитаемые острова», который также классифицируется Советом Европы как биогенетический заповедник.
Они являются частью Пустынных островов, Островного этажа, Пустыни Гранде и Бугио. В административном отношении они входят в состав прихода Санта-Крус, муниципалитета Санта-Крус, автономного района Мадейры.
4. Дикие острова
Острова архипелага Мадейра: Дикие острова — Изображение 5
Острова Сельвагенс — субархипелаг архипелага Мадейра, Португалия, расположенный в Атлантическом океане.
